La vida
El
universo subalterno termodinámico llamado vida, es capaz de autorealizarse
mantenimiento y mantener la integridad de su estructura de información, más
allá de la estructura, orden y desafíos espacio-tiempo propio del ambiente
genético evolutivo y metabólico ecológico. Donde para éste último, su
estructura de información es más pobre. Sin embargo, esta red de reacciones
químicas del universo subalterno, o vida “simplemente”, parece un concepto que
pretende hacernos escapar de esta realidad física, evolutiva, de cálculo y conocimiento.
Aunque nuestro concepto de vida, no haya avanzado más allá de cuando éramos
niños, podemos sacar una conclusión parcial, al definir la vida como el don que
hace tender al universo a la conciencia de su ser. De igual manera cualquier
otro concepto de vida, lo aseguramos, está entre los límites del lenguaje
(poesía-matemáticas), como por ejemplo los siguientes: la vida es el resultado final del disfrute de la conciencia como formas caóticas de
inteligencia y emociones creativas. O si lo prefiere: es la pirámide de la conciencia que sólo sobre ella misma descansa su
futuro.
Pensar en la vida permite iniciar el
fascinante estudio de la biología enmarcada entre la tendencia al equilibrio químico y a la
complejidad dinámica de sus códigos, la vida es la posibilidad absoluta de
romper la soledad de la materia
inorgánica. Hay organismos que de alguna manera adquirieron variedades fascinantes
de habilidades que concretamente les permiten adaptarse a su entorno natural y
social. Nos parece adecuado llamar al diseño de la naturaleza con el
calificativo de sumamente creativo. Sin duda, no se trata de una conducta
meramente aleatoria, sino de un comportamiento que de inicio resulta difícil de
caracterizar. Ahora bien, los organismo
que han adquirido este intricado conjunto de habilidades altamente organizadas
y articuladas, guardan los secretos del curso de la vida como cierta cantidad y estructura de información directa que define su naturaleza. Para ejemplificar esta postura filosófica de ver la
vida como estructura de información, exponemos el término de desafío cognitivo,
desprendido de la moderna rama de la biología llamada ciencia cognitiva.
De manera innata la curiosidad en los
seres humanos se haya motivada para emprender la búsqueda de regularidades,
leyes naturales, sociales, propuestas políticas, técnicas,…, dado que el área broca del cerebro nos
impulsa a explorar
[1]
. Al
parecer, esta curiosidad hace emerger las habilidades cognitivas del cerebro
humano, surgen como resultado de una evolución genética (FOXP2) que complejizó la
anatomía del cerebro
[2]
. Con
todo y eso, tal curiosidad no puede provocar conocimiento de forma computacional
[3]
.
Y por consiguiente, el cerebro humano es por default curioso de la realidad geométrica
y simbólica que le circunda; sin embargo, esconde mucho, aún más allá de la idea
computacional del funcionalismo de la inteligencia artificial, que indica que
el cerebro observa al universo como estructuras de información
[4]
,
que dan la sensación de conocimiento para nuestra mente consciente. Por ejemplo,
las proteínas responden al ideal de partícula
[5]
y los genes al ideal de símbolo
[6]
.
Es decir, la materia viva es
al mismo tiempo espacio-forma y código; de igual manera son sistemas que
evolucionan al equilibrio y al caos simultáneamente. Es
relevante para modelar el cerebro humano en términos de desafíos cognitivos,
precisar qué es estructura de información, misma que se da forma mediante el
símbolo y la partícula, además, qué rol relevante juegan en el modelado de la
mente consciente. La mayor parte de la materia y de la cultura de los
seres vivos incluyendo el cerebro y la sociedad; ambos, continuamente están
siendo remplazados por división y reparación celular; tejidos, órganos, mitos,
leyes y conceptos respectivamente, sólo permaneciendo su estructura de
información. El símbolo6,
es la unidad de información que codifica y estructura la realidad lingüística,
cultural y biológica en términos de palabras código que comunican y heredan
estructuras de información. De acuerdo con Roger Penrose4, debemos
mirar más allá de la dicotomía cuerpo y mente, si queremos extender las posibilidades de libertad creativa que
la ciencia aspira como ideal permanente. La partícula5, de acuerdo con la física
estadística
[7]
,
es en cualquier objeto la unidad en que puede ser descompuesto por un sistema,
donde su tamaño infinitesimal permite reducirlas a coordenadas y su momento físico. La bioquímica del cerebro está
gobernada su funcionalidad por la geometría de proteomas, que en su doblado construyen un espacio-forma
de partículas cargadas eléctricamente llamadas epítopes conformacionales, los cuales determinan
el funcional biológico sobre una estructura de información llamada bioquímica.
El desafío cognitivo que implica estudiar la vida, es
sin duda alguna, la tarea que nos debe ocupar a la hora de estudiar la
biología, es el proceso más relevante del intelecto, lo advertimos a los
estudiantes, no como un asunto estrictamente pedagógico, sino, como un asunto
estrictamente humanista de ver la vida.
Es posible investigar la información
disponible para la vida, al hacerlo, en principio nos enfrentamos con un problema científico de
enormes desafíos cognitivos, conocimiento generado por las observaciones
objetivas profundamente organizadas que los científicos publican en modernas
bases de datos de revistas
indexadas y evaluadas por factor de impacto. Lo más notable de esta tarea es hacer
conciencia que en esta gran variedad de reportes científicos, existen
limitaciones en relación con los sistemas de explicación resultantes, el
contenido generado del conocimiento es una experiencia de aprendizaje riguroso,
sin duda el estudiante debe partir del hecho de que su lenguaje especializado
le permite un margen de maniobra intelectual explícito, para organizar el
conocimiento en sistemas alternativos de explicación, debería agregar más allá
de cantidad de lenguaje especializado, la experiencia de su uso en la
composición escrita de un sistema
de conocimiento, que le dote de la sensibilidad y curiosidad de valorar
la vida en una cartografía de conocimiento científico y técnico complejo. Entre
los conceptos o nociones que los científicos usan para elaborar conocimiento, los hay de muchos grados de complejidad, en función
diferenciadora y analítica, algunos nos permiten caracterizar la célula y otros nos permiten
establecer y hacer de su fenomenología un discurso de explicación con reglas internas de la práctica de la razón que dotan de rigor a la
investigación sobre la vida, que constituyen la práctica científica de la ciencia biológica.
Afirmamos que el concepto de vida no es un concepto científico, en nuestra humilde opinión es
un concepto filosófico que la epistemología hace
converger sobre todo intento de discutir científicamente la realidad y no sobre su
contenido. Hablar de elementos
constitutivos de la materia viva de nuestro universo conocido, nos conduce a
una discusión biológica inmutable de su existencia, sin embargo, el concepto de vida que está organizado en las
ciencias biologías, desde la genómica, transcriptómica, proteómica hasta la fenómica, esconde su
organización intrínseca, que es su desafío cognitivo, que es el intento de determinar su estructura
de información, que no creemos que la biología y la física actual estén
próximas a resolver en definitiva lo desconocido de esta estructura de
información que permita unificar un concepto de vida. El concepto de racionalista necesario para
explorar científicamente la vida, debe ser tomado en cuenta para que permita
adquirir experiencia, en el cómo los hombres iniciaron a oscuras su viaje a la
realidad, donde nuestra humilde perspectiva nos conduce a retomar los grandes
pensadores que dentro de limitaciones históricas se acercaron a conceptos,
ideas arriesgadas y reveladoras que de inicio ellos no podían tener una
conciencia plena. Creemos que todos los estudiantes pueden acercarse a la biología
moderna por sus propios pensamientos creativos, si intenta no simplemente
compararse con los grandes pensadores del pasado y en su lugar da paso a una
actitud honrada de preguntar dentro de un trabajo intelectual, en un esfuerzo
por alcanzar creativamente la realidad, si podemos superar ser una especie de
superficie receptora de información, que acumula y luego combina, es decir, la
biología como cualquier otra ciencia nos exige creatividad como sujetos lingüísticamente
hablando, una actitud de reinventarnos permanentemente, no como un asunto de
fuerza bruta combinatoria de datos, sino como un asunto de talento.
Los biólogos tienden a pensar en
la vida por lo que hace, en lugar de lo que es. Por ejemplo,
los virus están en la
frontera de la vida. Ellos pueden imaginarse a estos como autómatas que simulan muchas
de las propiedades de la vida, pero que no puede sobrevivir sin los
organizadores vivientes. Para tomar esta línea de pensamiento más allá, consideremos: los virus están
constituidos de las mismas proteínas y ácidos nucleicos que forman
lo que nosotros pensamos como la vida. Esto lo subrayamos porque nosotros no
podemos definir la vida en base a lo que la forma. La vida simplemente no es la
vida porque es hecha de proteínas y los ácidos nucleicos.
En el caso de
una definición basada en procesos podría ser más útil. Los organismos vivientes hacen
cosas que las cosas inanimadas no hacen. Ellos comen, excretan las basuras, y
hacen más de su propio tipo. Ellos tienden a reaccionar a los estímulos externos
modificando sus comportamientos o los funcionamientos interiores. El problema
son tales propiedades que se ponen en correlación con la vida, pero ellas no la
definen. Es más, se encuentran estas propiedades en otros sistemas, en los robots y programas de computadora,
existen sistemas que generan copias de ellos y modifica su propio
comportamiento en respuesta a los estímulos.
[8]
¿Los robots y programas de computadora pueden verse como vivientes?, la mayoría
contestaría negativo, pero esta respuesta viene del prejuicio parroquial en
lugar de la investigación
científica.
Este argumento
expone un problema fundamental sobre el cómo nosotros entendemos la “vida”. Es
decir, nuestra concepción de vida, inevitablemente, se limita por lo que
nosotros sabemos, y esto es completamente basado en lo que nosotros vemos
alrededor de nosotros en la Tierra. ¿Por qué la vida en la Tierra es hecha de
proteínas y basada en la química de compuestos de carbono para la solución ácuea?, ¿nosotros tendemos a
asumir que toda la vida debe hacerse así? A pesar de la existencia de
computadoras y robots que imitan algunos de los procesos de la vida, la
necesidad de saber no nos debe conducir a hacer de ésta una receta de cocina; y
a pesar de la existencia de virus que son formados por proteínas y ácidos
nucleicos convencionales, pero que no cumplen con todos los criterios de las
cosas vivientes superiores, estos podemos considerarlos en la frontera de los
vivientes y los inanimados.
Estos argumentos exponen el debate
popular de la existencia de vida en Marte como no pertinente. Científicos han
sugerido que Marte, algún día en su pasado, fue caluroso y bastante húmedo para
organizar la vida,
[9]
con
la implicación que la vida todavía podría existir allí. Pero esto descansa en
la asunción que la vida marciana es (o era) formada como la vida de la Tierra.
Porque la vida en la Tierra es todo lo que nosotros sabemos, esta asunción no
puede justificarse en condiciones científicas, usted nunca puede alcanzar estadísticamente una conclusión válida basada en
un tamaño de muestra unidad.
Las gotas
microscópicas dentro de los meteoritos de Marte podrían representar los fósiles de bacterias. Cosas así exigen
sólo ser basadas en las comparaciones con bacterias terrenales o sus fósiles, y
no hay ninguna razón para suponer que esa bacteria marciana o sus fósiles
necesitan que sean parecidas a algo en la Tierra.
Claramente, lo
que nosotros necesitamos es un acercamiento a la definición de vida que no sea
ninguna presa del prejuicio parroquial. Las físicas podrían proporcionar una
respuesta. Por lo que se refiere la termodinámica, la rama de la física relacionada con la conservación de la energía y
su traslado. Es una conservación de la energía a través de una cantidad
llamada entropía
[10]
que nos va a medir el orden y el desorden de la materia. Pensamientos así es la Segunda Ley de Termodinámica -la tendencia de todos los sistemas a aumentar su entropía, para alcanzar una
situación en que ninguna parte del Universo contiene más energía que cualquier
otra-. El volumen de energía puede expresarse de otras maneras, como el volumen del orden, organización, complejidad o información. La
teoría general de la evolución afirma que toda criatura viviente, incluso el hombre, es resultado de una
combinación fortuita de los átomos y las moléculas de una supuesta “sopa” primordial.
[11]
Evitar el
aumento de la entropía es en general imposible, es decir, una parte del universo podría esforzarse por mantener su propio
volumen de orden o información al gasto de otras partes. Se dice que semejante universo subalterno está en
condiciones termodinámicas, “lejos
del equilibrio”. Los organismos vivientes hacen ejemplos buenos de tales
universos subalternos. Los procesos que nosotros asociamos con la vida ocurren
lejos del equilibrio--la vida exige el mantenimiento activo de estructuras en
el desafío de entropía, sin mantenimiento se deterioraría y desaparecería--.
Por ejemplo, las membranas
celulares no son bolsas pasivas, son superficies activas que
constantemente trabajan para asegurar la integridad de los volúmenes al gasto
del ambiente. En cuanto ellas detengan el funcionamiento, la célula deja de existir.
[12]
En general, la
vida podría pensarse como un juego de fenómenos en que las colecciones de átomos forman agregaciones
temporales que cambian el número de miembros, organizándolos en sistemas lejos
del equilibrio; eso contiene más
información, orden y estructura que sus ambientes; y donde la posición
enérgica se mantiene por actividades generadas dentro del sistema y al gasto de orden, información y
estructura fuera del sistema. La implicación es que la entidad viviente
normalmente es (pero no necesariamente siempre) separada del resto del universo
por un límite discreto, una membrana
celular.
[13]
,
[14]
Esto parece por
fin un progreso. ¿Pero lo es?, nosotros hemos movido el problema no a una fase
que nos lleve más allá, igualando la vida con todas las formas de sistemas que
están lejos del equilibrio. Algunas reacciones químicas y otros productos de
los procesos físico, están
lejos del equilibrio y no nos conducen a los modelos de gran complejidad orgánica. Incluso un montón de arena, o nieve antes de un
alud, podría pensarse de esta manera. Uno podría extender la definición de vida
incluso para abarcar el Universo entero, el pensamiento es visto como un sistema lejos del equilibrio. Hace el
orden que nosotros vemos en las estrellas y galaxias, el
reflejo de una propiedad del Universo que permite las desviaciones del
equilibrio. Están acostumbrados los físicos quánticos a imaginar las desviaciones pequeñas y efímeras
de las leyes de conservación de masa y energía, en la forma de la creación y
destrucción de partículas.
Hace la existencia en el mismo Universo de cosas vivientes--las desviaciones
más grandes, análogas-- ¿nos dice algo sobre la cara del Universo en que
nosotros vivimos? En ese caso, entonces la vida es una parte íntegra del Universo, tanto una parte del
Universo como la constante de Hubble y la curvatura de espacio-tiempo.
[15]
Como tal, buscar definiciones de vida como un fenómeno discreto es muy difícil, algunos dirían
en vano, pero sin duda es una causa sustantiva para el descanso del alma humana.
No hay ninguna respuesta simple a la pregunta de lo ¿qué es la vida?, eso no
incluye algún límite arbitrario. Sin semejante límite, cualquiera está vivo o
todo es nada. 11,
[16]
,
[17]
“Se
aproxima una crisis de
percepción. La complejidad del mundo ha llevado al ser humano a
simplificar la realidad, a abstraer la naturaleza para hacerla cognoscible y,
tristemente, a caer en la trampa de la dualidad. Bien y mal; objetivo y
subjetivo; arriba y abajo. Pero la tendencia a ordenarlo todo lo choca con la
misma realidad, irregular y discontinua. Muchos científicos ya han renunciado a
la ilusión del orden para dedicarse al estudio del caos, que acepta al mundo
tal y como es: una imprevisible totalidad.”
[18]
La aparente diversidad biológica en la tierra sólo existe si
lo comparamos con diferentes especies separadas de sus entornos, significa que las especies que coexisten
tienen que ser idénticas en términos de interacciones energéticas. Considere la
posibilidad de la biosfera como una red de reacciones químicas. Esto lleva a la conclusión de que las
fuerzas que impulsan la evolución de la vida se pueden encontrar dentro del
proceso de las transformaciones de las moléculas. Desde el punto de vista
termodinámico el sistema de partículas que reaccionan alcanzará un estado de
equilibrio cuando el producto
químico y los potenciales de los reactivos se igualan. Las interacciones
entre los compuestos químicos que participan en las reacciones concurrentes y el equilibrio de los potenciales químicos son la
esencia de la evolución biológica y su único motor. La evolución biológica no significa que las especies
se adaptan más a su medio ambiente debido al proceso de selección natural
[19]
. Desde el punto de vista físico de la vida en el planeta es
una red de reacciones químicas. La evolución sólo significa que todos los potenciales electro-químicos en la biósfera tienden
a ser iguales
[20]
. No es más útil y más "inteligente" que una
reacción química en un tubo de ensayo. No importa lo que la biósfera parece,
sorprendentemente para nosotros es simplemente el resultado de un gran
experimento químico de las reacciones concurrentes.
[21]
Arquitectura quiral de la vida
La homoquiralidad es un término usado para referirse
a un grupo de moléculas que se producen en espejo o quirales (ver figura 1), sin embargo, lo
sorprendente es que la mayoría de las biomoléculas se producen en espejo, o
quirales, las imágenes de cada una existen. ¿La vida es homoquiral?: las
proteínas contienen casi exclusivamente arreglos de aminoácidos en forma-L,
mientras que sólo biológicamente los azúcares que aparecen en el ARN y el ADN
tiene la forma-D. El mecanismo detrás de esta asimetría fundamental de la vida
sigue siendo un problema abierto. Investigadores sostienen que la
homoquiralidad de la vida es resultado de la simetría secuencial quiral
provocada por los acontecimientos del medio ambiente, ampliando así la teoría del equilibrio puntuado al
reino prebiótico.
[22]
En este caso, proponemos que la
homoquiralidad de la vida puede ser explicada por el paso de grandes tiempos
prebióticos, la hipótesis de equilibrio puntuado de Eldredge y Gould, por la cual el efecto quiral se produjo a
través de la alternancia de ciclos de estancamiento y de la intensa actividad
impulsada por cambios externos
[23]
.
Estamos tomando prestado el concepto de equilibrio puntuado como: la red de las reacciones químicas
libres, es un sistema de no-equilibrio abierto capaz de intercambiar energía
con el medio ambiente.
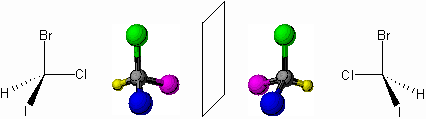
Fig. 1. Moléculas
quirales
Los funcionales tridimensionales de
las biomoléculas encuentran respuestas en un mundo tridimensional, se han
propuesto argumentos
antrópicos de forma independiente, por los filósofos y los científicos
para explicar por qué percibimos un universo tridimensional. El gran físico
Stephen Hawking en su obra monumental “La historia
del Tiempo” dice para el origen y la forma del universo: “vemos el universo en
la forma que es porque nosotros
existimos”
[24]
. Es
un buen ejemplo de un argumento antrópico.
La conjetura de Kant
[25]
de que la tridimensionalidad del espacio puede, de alguna manera, estar
relacionada con la ley del cuadrado inverso de Newton de la gravitación, fue el
primer paso en esta dirección. Su contribución tiene el mérito mismo que
sugiere que el problema de la dimensionalidad también puede ser tratado en el marco de la Física y no pertenece
exclusivamente al dominio de las matemáticas, ni a la de la especulación
filosófica pura. Una comprensión más profunda de la conjetura de Kant, tuvo que
esperar al surgimiento de la teoría
de campos. Se han encontrado que la estabilidad del sistema planetario,
molecular y atómico guarda una relación similar de estabilidad mecánica
[26]
dentro de un espacio tridimensional descrito por las ecuaciones de Poissonusadas
como concepto antrópico.
Es claro que la estructura del espacio físico es algo dado o independiente del
hombre, sin embargo, al final es una función de nuestro esquema conceptual.
Pero Bertrand Russell, supone que "la limitación de las dimensiones a tres
(...) es empírica."
[27]
La propiedad topológica tridimensional del universo está presente en los seres
vivos, no solamente como forma, sino fundamentalmente como funcionales
bioquímicos de la vida dados por las proteínas, porque las observamos así
[28]
.
Si los fenómenos naturales no ocurren separados del espacio-tiempo, podemos
decir, que tampoco se genera conocimiento científico separado del lenguaje-experiencia. Por
principio antrópico podemos hacer la suposición de que el primer universo
necesariamente debió contener aminoácidos.
La vida como rango de tiempo finito
La mayoría de los datos
demográficos indican un aumento casi exponencial de la mortalidad de adultos
con la edad, un fenómeno que ha sido explicado en términos de una disminución
en la fuerza de la selección natural actuando sobre la mortalidad por edad. Por
máxima verosimilitud los hallazgos
demográficos sugieren la existencia de muerte en edades avanzadas en los seres
humanos y los insectos dípteros, aparentemente en contradicción con los datos
de la teoría evolutiva, tanto por la pleiotropía
antagónica y la acumulación de mutaciones como la
conducción de la población por mecanismos genéticos.
[29]
En la investigación sobre
el envejecimiento se ha empleado por mucho tiempo, los modelos demográficos de
edad, la mortalidad de adultos específica se basa en un aumento de aproximación
exponencial de la mortalidad. El más conocido modelo es el de Gompertz
[30]
:
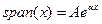
Dónde el envejecimiento depende de
las tasas de mortalidad, span(x); es determinada por dos parámetros, A y
α, que afectan independientemente y dependientemente la tasa de
mortalidad respectivamente. El patrón
Gompertzian de la tasa de aumento de la
mortalidad ha sido explicado por los biólogos evolucionistas en términos de una
disminución en la fuerza de la
selección natural
[31]
,
por ejemplo, la esperanza de vida humana en los países desarrollados ha
aumentado de manera constante por más de 150 años, a través de mejoras en la
salud pública y estilo de vida. Más personas por lo tanto viven lo suficiente
para sufrir la pérdida de funciones relacionadas con la edad y la enfermedad, hace que haya una necesidad de
mejorar la salud de las personas mayores. El envejecimiento es un proceso complejo de la
acumulación de daños, y ha sido considerado como experimental y médicamente
intratable. Esta visión ha sido reforzada por la constatación de que el
envejecimiento es un rasgo desfavorable, que evoluciona como un efecto
secundario de la acumulación de mutaciones o un beneficio para los jóvenes,
debido a la disminución en la fuerza de la selección natural en edades
posteriores. Sin embargo, importantes descubrimientos recientes muestran que
las mutaciones en los genes individuales pueden extender la vida útil de los
organismos en modelos de laboratorio y que los mecanismos implicados se
conservan a través de grandes distancias evolutivas, incluyendo a los
mamíferos. Estas mutaciones pueden mantener a los animales y la patología
funcional, libre a edades más avanzadas, y pueden proteger contra el
envejecimiento específico relacionado con las enfermedades, incluyendo enfermedades neurodegenerativas y el cáncer. Las indicaciones
preliminares sugieren que estos nuevos resultados del laboratorio pueden
también aplicarse a los seres humanos
[32]
.
La traducción de estos descubrimientos en tratamientos médicos plantea nuevos
retos, incluyendo el cambio hacia el pensamiento clínico de amplio espectro, la
medicina preventiva y la búsqueda de nuevas rutas para el desarrollo de drogas.
Las primeras evidencias de estudios de población de asociación genética también
han comenzado a implicar a la vía genotipo FOXO3A en la determinación
del rango de vida humana
[33]
.
Para una población objeto que está cerca de alcanzar el equilibrio
demográfico de selección débil, la fuerza de selección natural a edad t actúa sobre los cambios
proporcionalmente uniforme en la
probabilidad de supervivencia, y está dada por:
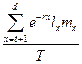
Dónde d es la última edad de reproducción, T es el tiempo de generación, mx y lx son respectivamente
la fecundidad específica y la
probabilidad de supervivencia, y r es la tasa intrínseca de crecimiento de la población. La dependencia del
envejecimiento sobre la fuerza de selección natural ha sido corroborada varias
veces, en especial en la mosca de la
fruta Drosophilia
[34]
.
En particular, se ha demostrado que el envejecimiento evoluciona rápidamente
cuando la fuerza de selección natural es
mayor a edades más avanzadas
[35]
.
Cuando la "barrera heterótrofo" fue finalmente rebasada en el precámbrico tardío, protistas herbívoros y
carnívoros surgieron casi al mismo tiempo, los dos grupos sin grandes
diferencias biológicas por separado
[36]
,
[37]
. La historia de la vida, se forma de muchos de estos casos.
La teoría
metabólica de la ecología formulada recientemente, tiene profundas
implicaciones para la evolución de las historias de vida
[38]
.
La tasa metabólica limita la expansión de la producción con
la masa corporal, de manera que los organismos más grandes tienen
tasas más bajas de la producción en masa base específica que los más pequeños. Las
implicaciones de esta limitación para la vida, es la evolución de su historia.
Se muestra que para una serie de historias de vida muy simples, la aptitud
darwiniana es igual a la tasa de natalidad menos la tasa de mortalidad. Así, la
selección natural maximiza tasas de natalidad y la producción reduce al mínimo
las tasas de mortalidad. Esto implica que la disminución del tamaño corporal en
general, se verá favorecida, ya que aumenta la producción, en tanto que la
mortalidad se ve afectada. Alternativamente, el aumento de tamaño del cuerpo se
favorece sólo si disminuye la mortalidad o aumenta el éxito reproductivo de
manera suficiente para anular la restricción de producción preexistente.
Adaptaciones que pueden favorecer la evolución de mayor tamaño, incluyen
cambios de nicho que disminuyen la mortalidad al escapar de la depredación o porque
la fecundidad aumentó en la explotación de nuevas fuentes de alimento
abundante. Estos principios pueden ser generalizados para comprender mejor la
relación íntima entre la moneda genética de la evolución y la moneda metabólica
de la ecología.
La teoría
metabólica de la ecología prevé que las consecuencias de muchos de los
atributos de los individuos, poblaciones, comunidades y ecosistemas deberían
ser relativamente sencillas calcularse a partir de los procesos metabólicos de
los organismos pertinentes. En concreto, muchos procesos de velocidad se deben
a la escala con el tamaño corporal y la temperatura, de la misma manera como la
tasa de masa metabólica específica surge:
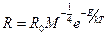
Dónde R es la tasa de algún proceso
biológico, R0 es una constante de
normalización, la M -1 / 4 término que da la función de poder
dependiente sobre la masa del cuerpo M,
y la exp --E / kT término o factor de Boltzmann da la dependencia de la temperatura exponencial
en términos de energía, una "activación", E, la constante de Boltzmann, k, y la temperatura, T,
en grados Kelvin. Esta relación es muy general, tanto internalista como entre
las especies. Se puede aplicar a la tasa de producción de biomasa por unidad de
masa, que se prevé ampliar a medida que es M -1 / 4 . Sí asumimos la temperatura constante, el factor de Boltzmann es una constante y
si tomamos el logaritmo de la ecuación anterior:
log(
tasa de producción masa-específica) = constante -1/4 log(masa del cuerpo)
La tasa de producción
masa-específica alométrica (la alometría
se refiere a los cambios de dimensión relativa de las partes corporales
correlacionados con los cambios en el tamaño total) es una disminución de la
función de la línea recta de la masa corporal, como se muestra esquemáticamente
en la línea en negro sólido de la figura 2.
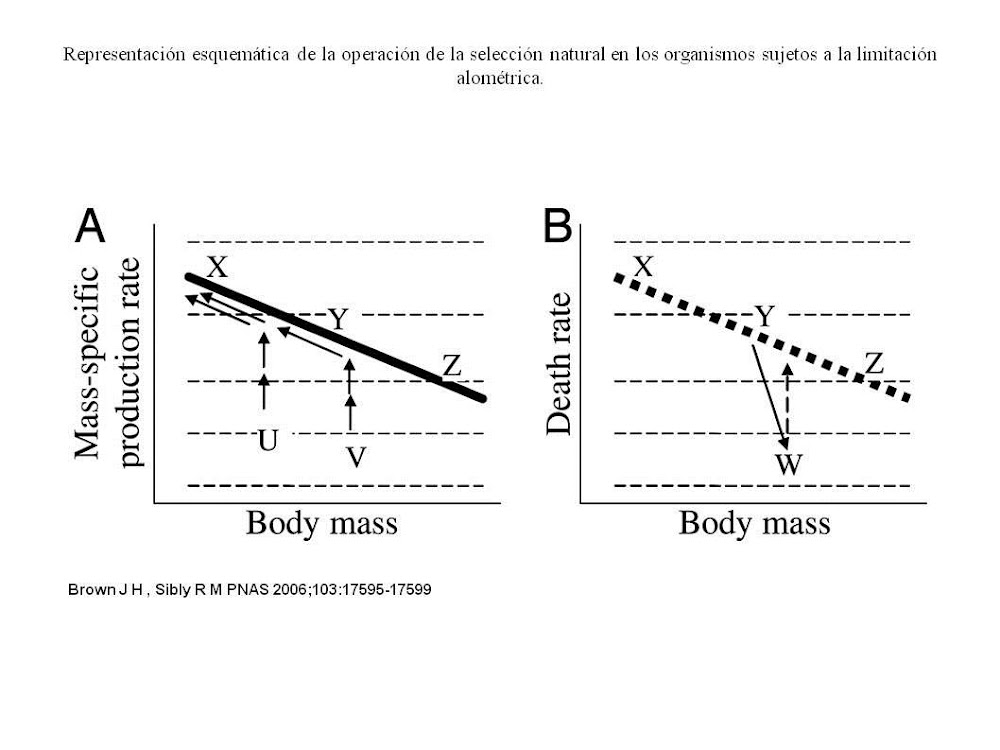
Fig.
2. Tasa de producción masa-específica alométrica.
Esto lo interpretamos: se
trata de una restricción de causalidad fundamental que limitan las opciones de
la historia de la vida como lo sugiere la teoría metabólica. El resultado es
que los organismos más pequeños tienen más recursos, en relación a la masa
corporal para asignar a la reproducción; por lo que al producir nueva biomasa,
nacen más individuos, y los genes se recombinan a un ritmo más rápido. ¿Cómo
funciona esta característica fundamental de la escala de producción alométrica y cómo afectan a la
historia de la vida en la evolución?
Poner la vida en la
teoría de la historia y la teoría metabólica juntas, de esta manera ofrece
ideas sobre algunos de los procesos más fundamentales de la biología de
organismos, la evolución y la ecología. En particular, se aclara la relación
íntima entre la moneda genética de la evolución y la moneda metabólica de la
ecología. La selección natural maximiza el estado
físico,
que los biólogos evolutivos han medido tradicionalmente en términos de las tasas de crecimiento diferencial de los alelos o tasas diferenciales
de la producción de crías descendientes con genes para un rasgo en particular.
Pero los hijos que llevan los genes están hechos de carne y hueso. Su
producción está alimentada por el metabolismo, y la tasa de la producción
obedece a las limitaciones de la alometría, la dependencia
de la temperatura, estequiometría, y otros procesos que
determinan la tasa metabólica
[39]
.
Elaboramos el concepto de vida con la teoría convencional de la historia para
aclarar la relación entre la tasa de producción y de las otras tasas vitales.
Entonces, utilizamos la escala alométrica de la tasa de producción para
analizar la evolución del tamaño corporal. Se muestra cómo éste limita la
alometría de natalidad y mortalidad, y cómo éstos a su vez limitan la evolución
del tamaño más pequeño o más grande.
La vida es su dinámica entre el estado de
equilibrio y caótico
El efecto estadístico de la irradiación interna
que proviene de las sustancias radiactivas presentes en los alimentos, en el agua y en el aire, las cuales, al ser ingeridas o
inhaladas comprometen la salud; en investigaciones
sobre la supervivencia de la raza Beagle (es una raza canina de caza
originaria de Europa), esta irradiación interna es
similar a los efectos del envejecimiento, con la excepción de que la muerte se
produce antes. Puesto que los procesos biológicos están a punto de equilibrio en la mayoría del tiempo, la observación anterior sugiere
que se produce la muerte cuando el tipo de salto ha superado la tasa de recuperación
medida suficiente, es decir, que las reservas están agotadas y el estado de
equilibrio no puede mantenerse. La teoría del estado estacionario de las tasas de mutación se deriva de los primeros
principios, y, a través de la teoría de tipo absoluto, es completamente general
y no se limita a un determinado tipo de alteración celular. Sin embargo, la
naturaleza de las alteraciones celulares se considera que puede conducir a la
muerte o a una ventaja evolutiva; los sistemas cada vez más jóvenes también se consideran
en el contexto de esta teoría
[40]
.
La teoría del estado
estacionario de las mutaciones fue desarrollada y aplicada a los numerosos
datos sobre el efecto de la radiación en los Beagles adquiridos durante veinte años. La teoría se utiliza para interpretar los datos
de HB Dorn sobre la incidencia de 21 tipos de cáncer
en los estadounidenses, tanto hombres como mujeres. La teoría muestra la
naturaleza de la heterogeneidad en la población de diversos trastornos. El
acuerdo encontrado confirma la teoría del estado estacionario de las mutaciones
de una manera sorprendente
[41]
.
¿Desechar nuestros
residuos afecta nuestro rango de vida? El envejecimiento biológico se debe a
varias causas, incluyendo el deterioro de las células, de las
mutaciones, la destrucción de las células de órganos a través de
infecciones, el envejecimiento por el hecho de no eliminar los residuos
nocivos, el envenenamiento de fuentes externas, y por el daño por radiación.
Todos estos procesos conducen a ecuaciones del tipo observado experimentalmente
para el proceso de envejecimiento, de modo que una evaluación adecuada de los
diversos factores que contribuyen al envejecimiento es especialmente
complicada. Muchos de los procesos no biológicos en los que los productos de
velocidad de reacción a la iniciativa de reacción de envejecimiento coinciden
con la ecuación de la tasa de envejecimiento. La corrosión de los metales es
exhibido como un ejemplo típico de éste y muchos otros ejemplos podrían darse
[42]
.
La ecuación general para la profundidad de la anestesia o modelo de GARD,
provocada por materias solubles en lípidos, este efecto puede ser imitado por
la acumulación de productos de desecho y se muestra para simular la curva de
envejecimiento típico, es bien sabido que los productos de desecho son una
causa frecuente de muerte.
El modelo de GARD es basado en
simulaciones de cómputo
[43]
,
utilizando el algoritmo de Gillespie
[44]
.
Este modelo es utilizado para las reacciones químicas y destinado a
proporcionar una herramienta
cuantitativa para el análisis detallado de la herencia sin la
información de transporte de los polímeros. Se trata de cambios discretos
estocásticos en los enlaces covalentes dictados por la ecuación diferencial:
[1]
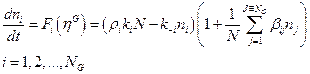
Dónde 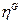 es un vector;
es un vector; 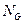 es el repertorio molecular de los compuestos prebióticos en el medio ambiente disponible;
es el repertorio molecular de los compuestos prebióticos en el medio ambiente disponible; 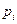 es la concentración externa de especies moleculares i;
es la concentración externa de especies moleculares i; 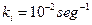 y
y 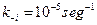 son catalizadas hacia adelante y hacia atrás las constantes de velocidad que se supone igual para todas las moléculas de simplicidad [que difieren en sus propiedades para el mejoramiento mutuo de la tasa];
son catalizadas hacia adelante y hacia atrás las constantes de velocidad que se supone igual para todas las moléculas de simplicidad [que difieren en sus propiedades para el mejoramiento mutuo de la tasa]; 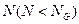 es el tamaño dado por los enlaces
es el tamaño dado por los enlaces 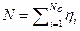 . Con
. Con 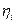 indicando el número de especies moleculares i
indicando el número de especies moleculares i
(es decir, la cuenta interna molecular de vectores 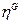 son
son 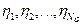 y
y 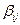 es un elemento de la
es un elemento de la 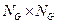 matriz positiva que define la red de interacciones mutuamente catalizadss y reguladss por un formalismo estadísticos. Teniendo en cuenta dos enlaces de composición
matriz positiva que define la red de interacciones mutuamente catalizadss y reguladss por un formalismo estadísticos. Teniendo en cuenta dos enlaces de composición 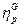 y
y 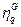 , su grado de similitud se define como el producto escalar:
, su grado de similitud se define como el producto escalar:
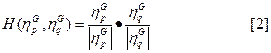
Donde 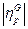 y
y 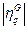 son normas euclidianas (H=1 representa la similitud perfecta, y H=0 indica ortogonalidad). La razón para suponer N < NG es que la transferencia de información se convierte en trivial para los conjuntos de gran tamaño.
son normas euclidianas (H=1 representa la similitud perfecta, y H=0 indica ortogonalidad). La razón para suponer N < NG es que la transferencia de información se convierte en trivial para los conjuntos de gran tamaño.
La dificultad en el estudio de la dinámica determinista del crecimiento de la motivación proceso de división en el modelo de GARD, es que, en principio, uno se enfrenta con una gran variedad de composiciones posibles de cualquier tamaño, desde un repertorio de NG: las moléculas del medio ambiente disponibles. Por lo tanto, si nos limitamos a una pequeña colección de NG =10 distintas especies moleculares y conjuntos considerados de tamaño 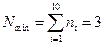 que se
que se
les permitió crecer a raíz de la ecuación (1) hasta que su tamaño alcanzado 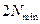 , tras lo cual se dividen exactamente en dos mitades. Sin embargo, hacemos hincapié en que la construcción matemática presentada aquí puede en principio con los enlaces de cualquier tamaño. En resumen, estamos tratando aquí de forma explícita con un espacio de composición en el que por cualquier enlace
, tras lo cual se dividen exactamente en dos mitades. Sin embargo, hacemos hincapié en que la construcción matemática presentada aquí puede en principio con los enlaces de cualquier tamaño. En resumen, estamos tratando aquí de forma explícita con un espacio de composición en el que por cualquier enlace 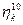 con el tamaño inicial
con el tamaño inicial 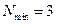 todos los enlaces hijos son accesibles del mismo tamaño que se calculan. Las tasas de mortalidad no se han incorporado y los conjuntos vacíos se evitaron al permitir la división de los enlaces en dos descendientes de igual tamaño, pero de otra manera la composición aleatoria (muestreo sin reemplazo) dificultan las conclusiones que pueden obtenerse por el análisis del sistema.
todos los enlaces hijos son accesibles del mismo tamaño que se calculan. Las tasas de mortalidad no se han incorporado y los conjuntos vacíos se evitaron al permitir la división de los enlaces en dos descendientes de igual tamaño, pero de otra manera la composición aleatoria (muestreo sin reemplazo) dificultan las conclusiones que pueden obtenerse por el análisis del sistema.
Un problema
importante es lo que la química más complicada podría hacer a estos sistemas no
macromoleculares con potencial hereditario. Creemos que la verdadera cuestión
es la de la organización de las redes químicas. Si no puede haber en
el mismo entorno distintas alternativas de organización de ciclos de redes autocatalíticas. Es notable que en 1971 Eigen descartó la autocatalítica de grupos de proteínas debido a la falta de herencia, es decir,
una proteína mutante introducida por casualidad (por un error de producción) no
pueda reproducirse sistemáticamente y se
pierde; mientras que un polinucleótido mutante siempre se puede replicar en el
mutante plantilla
[45]
.
Ahora nos sentimos obligados a abandonar la herencia de composición como una
forma de saltar hacia las unidades reales de la evolución. Hogeweg
[46]
distingue entre atractor y en el
almacenamiento basado en la herencia, en esta última
categoría se refiere claramente a los sistemas de base genética. Estamos de
acuerdo en que esta distinción es crucial en el análisis de los sistemas cuasi
biológicos. La esencia de los ácidos nucleídos desde el punto
de vista de la herencia es exactamente lo que pueden almacenar en una gran
cantidad de información, a la energía aproximadamente equivalente a niveles de
estabilidad, exactamente la propiedad que requiere de "almacenamiento". Un atractor de la información, en los sistemas
depende fundamentalmente del número limitado de estados estables alternativos
de enlace, como lo demuestra el análisis del modelo de GARD y es fundamental
para activar los mecanismos de desechos químicos.
La dinámica de división celular
La aparición de
un ciclo de la división celular no es una distribución exponencial: las
diferencias en los tiempos de generación de células hermanas se pueden derivar
de la teoría de la supervivencia de las poblaciones. La teoría de la
supervivencia Eyring Stover se ha aplicado a la cinética de la distribución de los intervalos de intermitóticas de células de mamíferos y, por inferencia a
la transición de la fase G1 en la síntesis de ADN (fase S)
[47]
.
La teoría se ajusta fielmente a los datos experimentales adquiridos por el tiempo cinemicrográfico, lapso de clonación de células HeLa en cultivo de tejidos, y
también sugiere la existencia de una sustancia iniciador lábil que interviene
en la fase de transición G1-S
[48]
.
La
posición alternativa plantea modelos del ciclo de la célula que permiten tener varias
transiciones al azar y la división celular asimétrica puede presentar una
propiedad que ha sido utilizada para apoyar el modelo de probabilidad de
transición del ciclo celular: que el valor absoluto de la diferencia entre las
hermanas entre los tiempos de mitosis celular que varía de una pareja de
hermanas a otra, y es descrita por una distribución estadística exponencial. Esta propiedad describe
que cada una de ellas postula la existencia de objetos que se dividen entre las
células hijas durante la división celular y cuyo número influye en la duración
del ciclo celular posterior, por ejemplo, los receptores de superficie de los
factores de crecimiento o complejos de transcripción que se realizan por
cromáticas hermanas. En el primer modelo, células hermanas reciben un número
idéntico de los objetos, que se utilizan para realizar múltiples transiciones
al azar que forman parte del ciclo celular. El segundo modelo es como el
primero, excepto que la compartimentación de los objetos entre el recién
formado y las células de hermanas es aleatoria. El tercer modelo es también
como el primero, excepto que todos los objetos son pasados a una de las células
de las hermanas. Estos hallazgos muestran que los mecanismos generales que son
responsables de la dispersión del tiempo mitótico, es decir, la correlación
entre tiempos de generación y la diferencia aparentemente, es una distribución
exponencial entre tiempos hermanos de generación. Podría ser una combinación de
la división celular desigual, múltiples transiciones del ciclo celular al azar
y la heterogeneidad de las células en la mitosis.
[49]
1.3.
150 años de “El origen de las especies”, C. Darwin.

La
importancia del concepto evolución difícilmente puede ser exagerada si decimos
que impregna todas las ciencias. De hecho, en los más de 150 años que han
transcurrido desde la publicación del “El origen de las especies”
[50]
, la idea
original de Darwin de que la evolución tiene lugar a través de la descendencia
con modificaciones tramitadas por la selección natural se ha convertido en un concepto clave en muchas
ciencias. Así, hoy en día se puede hablar, por
supuesto, de la biología
evolutiva, pero también hay disciplinas
evolucionistas en economía, la psicología, la lingüística, o ciencia de la
computación, por citar unas pocas. La teoría de la evolución de Darwin se basa
en la idea de selección natural. La selección natural es el proceso favorable a
través del cual los rasgos hereditarios más comunes (estructura de información)
pasan en las sucesivas generaciones de una población. Para poder medir este
proceso en una forma matemática precisa, J.B.S. Haldane y Sewall Wright incorporan, la llamada síntesis
moderna de la evolución de la década de 1920, el concepto de estado físico. Ellos aplicaron las ideas teóricas de la población a la
descripción de la evolución y, en ese contexto, se define estado físico como el número
esperado de descendientes de un individuo que alcanza la edad adulta. De esta manera fueron capaces de
llegar a una bien definida medida de la adaptación de los individuos y especies a su medio
ambiente.

El modelo de estado físico de herencia, tiene otra variante, el estado físico de espacio forma o llamado
por los biólogos paisaje de
adecuación, es un concepto topológico
-fitness landscapes- muy
importante en un espacio métrico abstracto de evolución. Este modelo determina
la capacidad de los ecosistemas y del medio ambiente para tolerar una actividad
en particular. Es menester mencionar que este método de capacidad asimilativa
se basa totalmente en la ciencia y asume que ésta puede restaurar el equilibrio
ecológico y la salud ambiental.
La forma teoría matemática de la evolución más
simple, surge cuando uno asume que el estado físico de una especie que no depende
de la distribución de frecuencias de las distintas especies en la población, es
decir, sólo depende de factores que son intrínsecos a la especie o las
influencias del medio ambiente. Sewall Wright
formalizó esta idea en términos de estados espacio forma, 1930, y en un contexto R (diagrama:
herencia, medio ambiente y residuos).
Fisher demostró su famoso teorema, que indica que la media de estado físico de
una población es una función no decreciente del tiempo, que aumenta
proporcionalmente a la variabilidad. Desde entonces,
mucho trabajo se ha hecho en este tipo de
modelos
[51]
,
[52]
,
[53]
,
[54]
, los
referimos al lector para su revisión. El enfoque en términos de estado de forma es, sin embargo,
demasiado simple y, en general, es evidente que el estado de forma de una especie dependerá de la
composición de la población y por lo tanto el cambio como consecuencia que la población evoluciona. Si se quiere describir la evolución a este nivel, la herramienta de
referencia está en la constante teórica de evolución en la teoría de juegos. Presentada en la
biología por Maynard Smith
[55]
una exaltación de la teoría de
los juegos desarrollados originalmente para la economía
[56]
, se ha convertido desde
entonces en un marco unificador para otras disciplinas, como la sociología o la antropología
[57]
. La característica principal de este aparato
matemático es que permite hacer frente a la evolución en un estado forma dependiente de la frecuencia o en otras palabras, con
las interacciones estratégicas entre las entidades, personas, grupos, especies,
etcétera; es la teoría de juegos evolutiva, pues, el enfoque genérico, incluye dinámica evolutiva
[58]
y constantes, como un espacio-forma constante
en la selección. Algunos modelos computacionales fractales de tales espacios-forma
los pueden generar con Fractal Explorer 2
[59]
, ver figura 4.
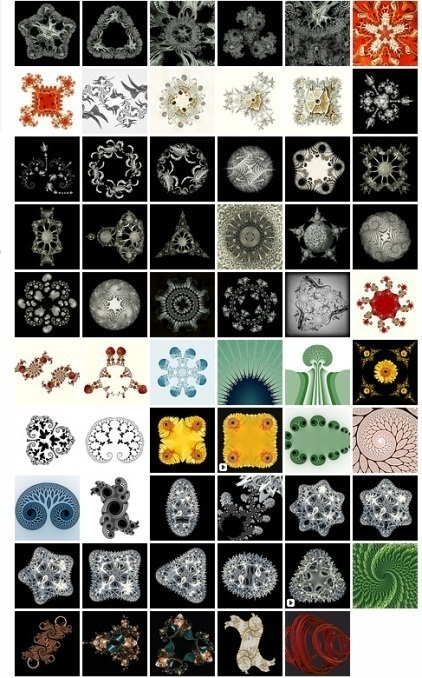
Fig. 4. Fractales, constantes espacio forma
(descargar animación http://www.vimeo.com/10531753 )
En su historia de treinta años, una gran cantidad
de investigaciones en la teoría evolutiva de juegos se ha centrado en las
propiedades y aplicaciones de la ecuación de réplica
[60]
. La ecuación de réplica fue introducida en 1978
por Taylor y Jonker
[61]
y se describe la evolución de las frecuencias de
tipos de población teniendo en cuenta su mutua influencia sobre su estado físico. Esta
importante propiedad permite que la ecuación de réplica pueda captar la esencia
de la selección y, entre otros resultados importantes,proporciona una conexión entre el concepto biológico
de las estrategias evolutivamente estables de Maynard Smith, con el concepto
económico de equilibrio de Nash62.
La ecuación de réplica se deriva en un marco
específico que involucra una serie de supuestos, comenzando con la de uno en el
finito, la población, bien mezclada y sin mutaciones. Por la población bien
mezclada, se entiende que cada individuo interactúa bien con los demás o por lo
menos uno tiene la misma probabilidad de interactuar con cualquier otro
individuo en la población. Esta hipótesis implica que cualquier individuo
interactúa eficazmente con un reproductor que utiliza la estrategia de medios
dentro de la población (un enfoque que se ha utilizado tradicionalmente en la
física bajo el nombre de aproximación de campo medio). Las desviaciones del
escenario de población bien mezclada, afectan fuertemente y no es trivial el
resultado de salida de evolución, de manera que es difícil para comprender en
principio. Estas desviaciones pueden surgir cuando se tiene en cuenta, por
ejemplo, el tamaño de poblaciones finitas, el aprendizaje alternativo/dinámica
de reproducción, o algún tipo de estructura (espacial o temporal como las
reglas sociales) en las interacciones entre los individuos.
1.4
Introducción a la teoría de la información en biología
La racionalidad científica clásica siempre ha
valorado, privilegiado, defendido y propugnado la objetividad del conocimiento,
el determinismo de los fenómenos, la experiencia sensible, la cuantificación
aleatoria de las medidas, la lógica formal aristotélica y la verificación
empírica. Pero la complejidad de las nuevas realidades emergentes durante este
siglo (genómica y proteómica), su fuerte interdependencia y sus interacciones
ocultas, por una parte, y, por la otra, el descubrimiento de la riqueza y
dotación insospechada de la capacidad creadora y de los procesos cognitivos del
cerebro humano, postulan una nueva conciencia y un paradigma de la racionalidad
acorde con ambos grupos de realidades.
Es deber de la ciencia ofrecer una
explicación rigurosa y completa de la complejidad de los hechos que componen el
mundo actual e idear teorías y modelos intelectualmente satisfactorios para
nuestra mente inquisitiva. Esto exigirá estructurar un paradigma epistémico que
coordine e integre, en un todo coherente y lógico, los principios o postulados
en que se apoyan los conocimientos que se presentan con fuerte solidez,
estabilidad y evidencia, ya sea que provengan de la filosofía, de la ciencia o
del arte. Pero la interdependencia de las realidades exigirá que este paradigma
vaya más allá de la multidisciplinariedad y llegue a
una verdadera interdisciplinariedad, lo cual constituirá un gran desafío para
la ciencia del siglo XXI.
Para 1900 Max Plack,
con su trabajo inicia la mecánica cuántica, pero es hasta la conferencia de Solvay 1927 que se enmarca de manera más formal la teoría
cuántica. Esta teoría constituye un radical rompimiento con la tradición física
anterior, porque aseguró que la naturaleza no se constituye fuera del saber.
Sin embargo, los fundadores de la teoría estipularon, cautamente que la teoría
no muestra la verdad total en este sentido, como una descripción construida
como sólo una manera de calcular acerca del futuro del conocimiento sobre información
básica provista de información pasada de un fenómeno natural.
La era moderna se creó probablemente
con la obra de Descartes, cuando este ilustre personaje separó la concepción
mente-materia para cualquier evento. Este movimiento liberó a la ciencia de los
dogmas religiosos y del constreñimiento de los primeros años, y les permitió a
los científicos introducirse en las más importantes regularidades matemáticas
del mundo físico observado. El propio
Descartes concede que la interacción mente y materia ocurre dentro de los
confines de un cerebro humano, pero el carácter determinístico del mundo físico
especificado por la mecánica de Newton parecían gobernar completamente fuera de
la mente e incluso dentro de nuestros cerebros, cualquier interferencia de la
mente con las ideas del funcionamiento de la materia se descartaba. Así la idea
de un universo totalmente mecánico, controlado por leyes físicas universales,
se volvió el único dogma de la ciencia. Puede imaginarse rápidamente que dentro
del entorno dominado por tal pensamiento habría una fuerte oposición a las
demandas radicales de los fundadores de la teoría cuántica que señala que los
conocimientos humanos conscientes deben tomarse como la base de nuestra teoría
fundamental de la naturaleza.
Todavía la oposición a este cambio
profundo en el pensamiento fue menos feroz que lo que uno podría haber
supuesto. Pero en el extremo nadie discutió que el resto de las ciencias que
nosotros podemos explorar y la teoría cuántica en términos prácticos descansaron
en ángulo recto con el hecho de introducción de la innovación teórica. El
cambio filosófico fue importante porque reformuló la economía, la
tecnología y la vida social, a partir de
que se inculcó en las mentes de académicos y técnicos la teoría cuántica.
Los nuevos modelos del pensamiento y
cálculos que ellos engendraron trajo bellamente en la medida que aterrizaron
estos pensamientos en problemas prácticos específicos, una pregunta: ¿cómo
hacer para traer a las culturas esta visión de la naturaleza?
Hay algunos físicos descontentos con
el éxito práctico y quieren entender lo que el éxito práctico de estas reglas
computacionales están diciéndonos sobre el mundo en el que vivimos, los
esfuerzos por lograr semejante reto, actualmente han trastocado la economía, la
sociología, las artes y la actual revolución del conocimiento biológico.
El problema radical que nos ocupa
aquí reside en el hecho de que nuestro aparato conceptual biológico, el que
creemos riguroso —centrado en la objetividad, el principio de causalidad, el
determinismo, la experiencia, la lógica formal, la verificación—, resulta
corto, insuficiente e inadecuado para simbolizar o modelar realidades que se
nos han ido imponiendo la cultura, ya
sea en el mundo subatómico de la física, como en el de las ciencias de la vida
y en las ciencias sociales. Para representarlas adecuadamente necesitamos
conceptos muy distintos a los actuales y mucho más interrelacionados, capaces
de darnos explicaciones globales y unificadas.
Debido a esto, en las tres primeras
décadas del siglo XX, los físicos hacen una revolución de los conceptos
fundamentales de la física; esta revolución implica que las exigencias e
ideales positivistas no son sostenibles ni siquiera en la física: Einstein
relativiza los conceptos de espacio y de tiempo (no son absolutos, sino que
dependen del observador) e invierte gran parte de la física de Newton;
Heisenberg introduce el principio de indeterminación
o de incertidumbre (el observador afecta y cambia la realidad que estudia)
y acaba con la objetividad; Pauli formula el principio de exclusión (hay leyes-sistema que no son derivables de
las leyes de sus componentes) que nos ayuda a comprender la aparición de
fenómenos cualitativamente nuevos y nos da conceptos explicativos distintos,
característicos de niveles superiores de organización; Niels Bohr establece el
principio de complementariedad: puede haber dos explicaciones opuestas para los
mismos fenómenos físicos y, por extensión, quizá, para todo fenómeno; Max
Planck, Schrödinger y otros físicos, descubren, con la mecánica cuántica, un
conjunto de relaciones que gobiernan el mundo subatómico, similar al que Newton
descubrió para los grandes cuerpos, y afirman que la nueva física debe estudiar
la naturaleza de un numeroso grupo de entes que son inobservables, ya que la
realidad física ha tomado cualidades que están bastante alejadas de la
experiencia sensorial directa.
Por esto, el mismo Heisenberg dice
que "la realidad objetiva se ha evaporado" y que "lo que
nosotros observamos no es la naturaleza en sí, sino la naturaleza expuesta a
nuestro método de interrogación"
[62]
,
es decir, el sueño del hombre de una realidad determinista, dejó el paso al
caos de la naturaleza.
En el sentido racional de ver a los
datos empíricos de la naturaleza como fabricados fuera de la mente, se
defiende, que la experiencia de los últimos setenta años hacen pensar en la
racionalidad de tomar esta interpretación en serio: más en serio que los
fundadores de la teoría cuántica las tomó en su momento. Básicamente, ellos
dijeron, cautamente, que el formalismo matemático es una herramienta útil por
formar expectativas sobre nuestro saber futuro en base a nuestro pasado. Esa
demanda ha sido ahora abundantemente incorporada, también en campos muy lejos
del estrecho confín de las físicas atómicas. Esta revolución que trajo la
teoría cuántica, ahora está en estrecha relación con la teoría de la
información, que en mucho juntas son la proteómica, la genómica y otras muchas
áreas de la biología y la salud humana.
La entropía
e información a menudo están en conflicto en la literatura. Una
comprensión precisa matemática e intuitiva de la noción de información y su
relación con la entropía es crucial para las aplicaciones, por ejemplo, en la
biología molecular, y desde luego vital para un cambio del espíritu curricular
de formación en los laboratorios de Q.F.B., comencemos para aclarar este cambio
de paradigma de docente de laboratorio, por el concepto original de entropía dado por Shannon 1948 y de
principio es necesario distinguir los conceptos de partícula y símbolo.
Con la partícula es posible obtener
cambios conformacionales que afectan la función de las proteínas y mediante el
símbolo de un alfabeto polipéptido o nucleótido es posible obtener la información de mutaciones o la arquitectura de
diseño que constituyen en un proceso biológico la proteína.
La partícula es estudiada en su dinámica física por la termodinámica y la física estadística, mientras el símbolo es abordado en un
escenario descrito por la mecánica de códigos en
la promisoria área llamada teoría de la bioinformación.
Es importante entender la diferencia entre entropía en el ámbito de la teoría
de la información y en el de la termodinámica para comenzar adentrarnos en este campo del conocimiento relacionado fuertemente
con el diseño de nuevas drogas y pruebas de robustez mutacional, entre sus
principales aplicaciones.
Uso de incertidumbre de Shannon
La teoría de entropía de Shannon, es
una medida de incertidumbre sobre la identidad de los objetos de un conjunto.
Aunque suele usarse entropía e incertidumbre como términos intercambiables, éstas
nunca pueden decir información.
[63]
Hay una relación simple entre los
conceptos de entropía dentro de la teoría de la información y el de la
termodinámica de Boltzman-Gibbs. La
entropía de Shannon o incertidumbre se define con respecto a un observador
particular sobre el estado de
un sistema. El ejemplo más simple de un sistema de estados es una variable aleatoria -random- objeto matemático que puede
pensarse como un dado de N lados diferentes, es decir, la probabilidad de cualquier lado o estado de N tiene la misma posibilidad
de ocurrir para todos los estados de N.
En el campo biológico podemos pensar
convenientemente en un polímero de longitud fija (número fijo de monómeros) que pueden asumir un estado
cualquiera de n posibles estados donde cada posible sucesión
corresponde a un posible estado. Así para una sucesión hecha de monómeros de tamaño L de un alfabeto de tamaño D,
tendríamos N=DL. Diríamos que la incertidumbre
calculada describe la observación efectuada sobre la verdadera identidad de la
molécula (entre un número muy grande de moléculas preparadas idénticamente: conjunto o totalidad), dado que sólo tiene el
observador cierta cantidad de conocimiento
probabilístico. La molécula
hipotética juega el papel de una variable random, si tenemos dada su distribución de probabilidad -el
conjunto de probabilidades p1,…,pN para
encontrar sus posibles estados-. Esta
molécula random, la denotaremos con “X” que
contendrá los nombres x1,…,xN de sus N estados. Si X tiene xi estados con probabilidad pi,…,pN,
entonces la entropía H de X es dada por la fórmula de Shannon:
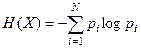
La base logarítmica corresponde a la unidad escogida para medir información. De manera análoga Shannon define la entropía de una función de distribución continua en una distribución de densidad:
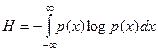
Con N dimensiones de distribución p(x1,…,xN)
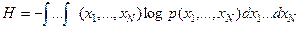
i fuera el caso de dos argumentos “x” y “y” la entropía de p(x,y) estaría dada por:
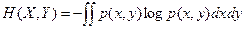
No hemos especificado hasta aquí la
base del logaritmo para las fórmulas anteriores. Especificando la base se
asigna unidades a la incertidumbre. A veces es conveniente usar el número de
posibles estados de X como la base del logaritmo (para el caso binario
cero-uno la base 2 es la conveniente). ¿Cómo aprendemos nosotros algo en la
vida de un sistema? Hay dos opciones: obtenemos la distribución de probabilidad
usando conocimiento previo (por ejemplo, tomando parte del sistema y obteniendo
teóricamente los demás estados por evolución) o haciendo medidas sobre él, esta
última vía no nos permitiría conocer todos los estados, situación que asume los
estados con la misma probabilidad. En ambos casos, la diferencia entre la
entropía máxima y la entropía restante después de que nosotros: o hemos hecho
nuestras mediciones o hemos examinado el sistema, es la cantidad de información
que nosotros tenemos sobre el sistema. La definición que nos conduce este
pensamiento es que la información
es una cantidad relativa. Mide la diferencia de incertidumbre, en el
caso anterior entropía, antes y después de la medición, y así nunca puede ser
absoluta, como lo es para el caso físico de energía potencial. De hecho, no es una analogía mala para referirse a la entropía como “la información potencial”,
porque potencialmente toda la entropía de un sistema puede transformarse en
información (por ejemplo por medición).
Si analizamos más profundamente la
ecuación de H(X) y además se mide en bits, una interpretación de
incertidumbre relacionada con los números más pequeños “
1,0”
o “si, no”, una pregunta necesaria sería sobre el promedio para identificar el
estado de una variable aleatoria X. Debido a que esta serie de signos
de interrogación si/no pueden pensarse como una descripción de la variable
aleatoria, la entropía H(X) también puede verse como la
longitud de la descripción más corta de X
[64]
.
En el caso en que nada es conocido acerca de X, esta entropía es dada por
,
que puede asumirse como el valor máximo de H(X).Esto
ocurre si todos los estados son igualmente probables: pi=1/N; i =1,…,N.
Si algo –más allá del número posible de estados N- es conocido sobre X,
esto reduce nuestro número necesario de interrogaciones, o la longitud de
medición necesaria para describir X. Si por ejemplo yo conozco que el
estado X= x7, es muy probable que mi incertidumbre sobre X va a ser más pequeña.
En el análisis anterior, la
información era la diferencia entre la máxima entropía y la entropía real del
sistema. En un sentido más general, la información mide la cantidad de correlación entre dos sistemas y
reduce la diferencia entre entropías en casos especiales. Para definir
propiamente información, nos permitiremos introducir otra variable aleatoria o
molécula llamada Y que puede estar en los estados y1,…,yM,
con probabilidades p1,…,pM.
Ahora podemos junto con la entropía H(Y) introducir la entropía de
juntura –intercepción-, H(XY) mide la incertidumbre en la juntura
–unión- del sistema XY –que puede estar en N
 M estados-. Si X y Y son variables aleatorias independientes –por ejemplo, dos dados que se tiran
independientemente- . La entropía de la juntura será justo la suma de las
entropías de cada una de las variables aleatorias. No para que se unan X y Y de algún modo. Por ejemplo, imagine dos monedas que se
pegan a una cara. Entonces la cabeza de una moneda siempre implicara las colas
–ir detrás de- para la otra y viceversa.
Encolándolas juntas, las dos monedas pueden asumir sólo dos estados, no cuatro y
la entropía de la unión es igual a la entropía de una de las monedas. Lo mismo
es válido por la unión de dos moléculas no ligadas al azar. Observe primero que
esas moléculas –DNA, proteínas- no se ligan al azar, en segundo lugar, el
ligado es efectuado por especificidad mutua que requiere de esa parte de la
secuencia de una de las moléculas actuando recíprocamente con la secuencia de
otra, para que la entropía de la juntura del par sea mucho menos que la
entropía de la sumas de cada una. Realmente este encadenamiento introduce
fuertes correlaciones entre los estados X y Y: si yo sé
el estado de una, yo puedo hacer predicciones fuertes sobre el estado de la
otra molécula. La información que una molécula contiene acerca de la otra es
dada por:
M estados-. Si X y Y son variables aleatorias independientes –por ejemplo, dos dados que se tiran
independientemente- . La entropía de la juntura será justo la suma de las
entropías de cada una de las variables aleatorias. No para que se unan X y Y de algún modo. Por ejemplo, imagine dos monedas que se
pegan a una cara. Entonces la cabeza de una moneda siempre implicara las colas
–ir detrás de- para la otra y viceversa.
Encolándolas juntas, las dos monedas pueden asumir sólo dos estados, no cuatro y
la entropía de la unión es igual a la entropía de una de las monedas. Lo mismo
es válido por la unión de dos moléculas no ligadas al azar. Observe primero que
esas moléculas –DNA, proteínas- no se ligan al azar, en segundo lugar, el
ligado es efectuado por especificidad mutua que requiere de esa parte de la
secuencia de una de las moléculas actuando recíprocamente con la secuencia de
otra, para que la entropía de la juntura del par sea mucho menos que la
entropía de la sumas de cada una. Realmente este encadenamiento introduce
fuertes correlaciones entre los estados X y Y: si yo sé
el estado de una, yo puedo hacer predicciones fuertes sobre el estado de la
otra molécula. La información que una molécula contiene acerca de la otra es
dada por:
I(X : Y)=H(X:Y)= H(X) + H(Y) – H(XY)
Es decir, la información es la suma
de las entropías de cada una, menos la entropía de la juntura. Las entrañas
entre X y Y en la notación para la
información estándar; se supone así, recuerde lector que la información es una
cantidad simétrica: lo que X sabe de Y, es la entropía condicional. Dicho de otra manera, la
entropía de X condicionada sobre Y, es la entropía de X dada Y,
esto se denota por H(X/Y) y se lee H de X dada Y, y se calcula como:
H(X/Y) = H(XY) –H(Y)
Esta fórmula es autoexplicativa: la
incertidumbre que yo tengo sobre X si Y es conocida, es exclusivamente
la incertidumbre sobre el sistema de la juntura menos la incertidumbre sobre Y. Si deseáramos conocer la entropía Y sin tomar en cuenta X, el concepto se llama entropía marginal. El concepto de entropía condicional podemos
rescribirlo:
I(X : Y)= H(X) – H(X/Y)
Ya hemos analizado el caso de las
variables independientes H(X/Y)= H(X) + H(Y), en el que la
información es una medida de
la desviación de independencia. De hecho, la cantidad de mediciones
exactas de la entropía de X o Y es reducida por el conocimiento de la otra variable
respectivamente.
Sí yo tengo un saber
 0 de las moléculas, me permite hacer predicciones más
exactas sobre la otra: esto es lo que queremos decir a
través de información en un lenguaje ordinario. Notar que esta
definición reduce en el ejemplo dado en líneas atrás –información como
diferencia entre entropías- si solo las correlaciones posibles están entre X y Y, mientras en la ausencia de la
otra molécula es equiprobable – significa que
cualquier sucesión es igualmente probable-. En este caso la entropía marginal H(X) debe ser máxima (H(X)=
log N) y la información es la diferencia entre la máxima y la real
entropía -es decir condicional-, como
dijimos antes.
0 de las moléculas, me permite hacer predicciones más
exactas sobre la otra: esto es lo que queremos decir a
través de información en un lenguaje ordinario. Notar que esta
definición reduce en el ejemplo dado en líneas atrás –información como
diferencia entre entropías- si solo las correlaciones posibles están entre X y Y, mientras en la ausencia de la
otra molécula es equiprobable – significa que
cualquier sucesión es igualmente probable-. En este caso la entropía marginal H(X) debe ser máxima (H(X)=
log N) y la información es la diferencia entre la máxima y la real
entropía -es decir condicional-, como
dijimos antes.
La entropía en la termodinámica
R. P. Feynman sigue presente, con sus
aportaciones al mundo de la física, vectores, grupos y espacios se agrupan
alrededor del concepto de partícula
[65]
,
que diremos de inicio de una manera breve, es diferente con la teoría de
Shannon basada en el símbolo
[66]
. Misma teoría que ahora describe la
complejidad de los genomas
[67]
.
Comentaremos que la entropía termodinámica de Boltzmann-Gibbs,
matemáticamente son muy similares sólo que la distribución de probabilidad pi es dada por la distribución Boltzmann de relevancia por el concepto de grados de libertad –posición
y cantidad de movimiento-:
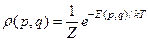
,
y la cantidad termodinámica se hace dimensional al multiplicar las
dimensiones de incertidumbre de Shannon por la constante de Boltzmann. Se dice
que no se puede medir todos los grados de libertad termodinámicos de forma
continua, porque es imposible que el aparato de medición pueda tener una
resolución infinita que pueda además afectar al sistema- dicho por Schrödinger-. Más importante
aún, el equilibrio termodinámico asume que todas las entropías de un sistema aislado están en su
máximo, así que no hay ninguna correlación en sistemas en equilibrio
termodinámico, y por consiguiente allí no hay información. Esto es
importante para nuestro propósito,
porque implica un corolario: la información contenida en los genomas biológicos
garantiza que la vida de un sistema está lejos del equilibrio termodinámico. La teoría de la información puede
verse así como un tipo de termodinámica lejos del equilibrio. No olvidar que definimos información como la
cantidad de correlación entre dos sistemas, como el instrumento que mide la cantidad de entropía entre dos sistemas y la información sobre la
que un sistema tiene de otro.
La información siempre es acerca de algo, si no puede especificar qué
información es ese algo, entonces estamos tratando con entropía y no con
información. Podríamos llamar a la entropía como el borde en una situación de
abuso del lenguaje: “información inútil”. Recordar que la discusión previa dar
entender que la información es sólo definida como relativa a un sistema de
información de algo, por consiguiente nunca es absoluta. En este punto es donde
el signo o símbolo se reconoce distinto del concepto teórico de partícula en la
física estadística.
Apoyándonos en la obra de David Deutsch, "El Tejido de la Realidad"
[68]
pretendemos explicar el campo de acción de la
ciencia biológica y sus ramas de estudio. David Deutsches un físico teórico
muy conocido cuya investigación se centra en la física cuántica y el campo
relativamente nuevo de la computación cuántica. Es miembro del Centro de
Computación Cuántica, que forma parte de la Universidad de Oxford.
De acuerdo con Deutsch la explicación de
nuestras teorías científicas actuales son la física cuántica, evolución, cálculo y conocimiento. Uno de los objetivos principales es mostrarnos
cómo estos cuatro temas están relacionados entre sí y nos permiten una
comprensión más profunda de "El Tejido de la Realidad" que cada uno
de estos por sí solo es capaz de permitir. Y continúa afirmando que la
combinación de nuestras mejores teorías en las cuatro áreas nos acerca a una
teoría del todo. Con esa teoría del todo, lo que no significa la búsqueda de
una teoría unificada de la física, que combinaría las teorías de la relatividad
general, la teoría cuántica, las fuerzas nucleares y el electromagnetismo en
una sola teoría. Se refiere a una teoría que nos da un profundo conocimiento de
nuestro mundo en una especie superior de nivel. Es sorprendente que Deutsch lo sugiera, ya que parece que esa teoría tendría
que ser reduccionista. Las teorías de la biología proporcionan explicaciones de la realidad
más que predecir el resultado de los experimentos. Siempre hemos considerado a
los biólogos para a hacer más describiendo la naturaleza que explicando la naturaleza.
Consideramos que la idea alternativa de que la biología no sólo describe y
predice la naturaleza, hace algo más, ya que proporciona una explicación más
profunda de por qué la naturaleza se comporta de cierta manera. Deutsch alega con razón que las modalidades específicas por
sí mismas no son importantes o interesantes, pero lo que es importante e
interesante es la comprensión de las leyes fundamentales que describen,
explican si se quiere, la aparición de estos patrones. Una vez hemos descrito
cómo la naturaleza se comporta, de preferencia con fórmulas matemáticas, ¿ya
está hecho? Es un hecho empírico que la naturaleza se comporta de manera
sistemática, que se puede describir con los modelos y fórmulas matemáticas. Y
nos parece que no es la verdadera belleza de la vida eso. Pero no tenemos
conocimiento de ninguna teoría que haga otra cosa que describir y predecir los
fenómenos en la naturaleza.
Un síntoma de Deutsch y otros en la búsqueda
de un significado más profundo detrás de los fenómenos de la vida es la idea de
una interpretación
cuántica. Es una idea posible
gracias a que el comportamiento de la naturaleza no se apega a la física como
una descripción en términos matemáticos. Basta decir aquí que su cadena de
razonamiento excluye la posibilidad de que el universo que nos rodea no
constituye la totalidad de la realidad. En conclusión, hay un sentido en el que
hay cosas tales como: ambientes Cantgotu
[69]
. Pero sólo si uno acepta las
definiciones más extremas de los entornos como estructuras de información.
Nosotros analizamos a la biología como fenómenos emergentes sobre las
que hay cosas para comprender que no pueden ser explicadas en base a teorías de
nivel inferior. Thomas Kuhn sostiene que la
ciencia es mucho más determinada por factores culturales, políticos y otros
subjetivos. Deutsch en su propia experiencia nos dice
una idea alterna de la cual simpatizamos más: la ciencia progresa de hecho
sobre la base de una mente abierta a nuevas teorías, la crítica objetiva, el
diálogo racional y la supervivencia de las mejores teorías. El argumento de Deutsch sugiere que la vida inteligente podría tener un
efecto muy grande en el universo, y por lo tanto no puede decirse que es
insignificante en términos del desarrollo a gran escala del universo. Su
argumento es que con la tecnología significativa, la vida inteligente podría
ser capaz de hacer grandes cosas, como el cambio del ciclo de vida de las
estrellas, o la vida de organismos con reparación infinita; solo es cuestión de
que se dé en el corazón de los hombres de ciencia una emoción tan fuerte que
proporcione la perseverancia para alcanzar por medio de la razón tal meta.
La coherencia
matemática como requisito para
hacer la biología está lejos de ser un criterio suficiente para decirnos si es
probable que estemos en las ideas correctas hacia la realidad. Sin embargo, la
coherencia matemática tiene la ventaja de que es claramente algo estético y al
mismo tiempo algo objetivo. Normalmente las condiciones físicas, químicas,
genéticas y geométricas de proteínas tienen más importancia en una observación
con el aparato matemático, por la posibilidad de confirmar o refutar
experimentalmente el índice real de su validez.
Nuestra comprensión de la realidad llamada vida está todavía muy lejos
de su objetivo, porque no sabemos cuánto es lejos o cerca. La biología moderna,
atendiendo al argumento de Deutsch sobre unificación
de las teorías de la física cuántica, evolución, cálculo y conocimiento; modifica fuertemente su arquitectura
para su estudio. Los nuevos campos de la biología están muy lejos de ser
reducibles a leyes físicas, pero no tenemos ninguna razón para creer que el
comportamiento biológico no sea, en su raíz, puramente dependiente de acciones
físicas. Hoy se acepta que la biología de forma crucial obedece a las reglas de
la mecánica
cuántica, como vaticinó Schrödinger en 1944 en su libro ¿qué es la vida?,
a partir de una dosis de humildad intelectual:
“Me parece inverosímil que nuestra comprensión del mundo represente una
etapa definitiva o final, un máximo o un óptimo desde cualquier punto de vista.
Con esto, no estoy queriendo decir simplemente que la continuación de nuestra
investigación en las diversas ciencias, nuestros estudios filosóficos e
intentos religiosos vaya a perfeccionar y mejorar nuestra presente perspectiva.
No hay ningún motivo para creer que nuestro cerebro sea el supremo nec plus ultra de un órgano de pensamiento en
el cual se refleja el mundo. Sería más razonable pensar que una especie pueda
adquirir un dispositivo semejante que guarde con el nuestro la misma relación
que el nuestro guarda con el de un perro o el de éste animal con el de una
babosa”.
[70]
Schrödinger
nota que un organismo vivo se mantiene sorprendentemente ordenado, a pesar de
que la agitación térmica y otros comportamientos estadísticos con ella
relacionados tienden a desordenar cualquier estructura constituida por muchas
partículas. Así, llegamos a una segunda pregunta: ¿cómo puede ser creado y
mantenido el orden (o la ordenación) de un ser vivo? Se trata de entender cómo,
por ejemplo, una célula huevo ya bastante ordenada y compleja, puede generar un
ser pluricelular, aún más ordenado y complejo. Schrödinger respondió esta
pregunta distinguiendo dos maneras de producirse orden, ambas de interés
biológico, que denominó “orden a partir del desorden” y “orden a partir del orden”. Y es que la materia viva, por la peculiar
organización de sus átomos en cristales aperiódicos, absorbe entropía negativa
del ambiente y se resiste a la degradación.
Lo más común es aceptar que una teoría biológica pertenece a la realidad
si guarda una fuerte relación apariencia-realidad. Schrödinger desafió esta
correlación de objetividad, en este mismo sentido, Nietzsche nos advierte que hay muchas maneras de hablar de
un evento, y que ninguna de ellas se acerca más que las otras al modo de ser
las cosas en sí mismas. Apariencia realidad, relación que muchos llaman verdad
en la ciencia de la naturaleza. Al igual que en un sentido poperiano, decimos
que no tenemos acceso a la verdad con los medios de la ciencia, sin embargo,
pensamos que las mejores teorías (creencias) no son las que más se justifican,
sino las más útiles para nuestra civilización, dado que no se conoce ninguna
teoría que sea inmune a la duda de su asertividad. Puesto que la verdad es una idea
absoluta y consiste su aproximación en una distancia vaga, la tarea científica
es humildemente una correspondencia lenguaje-realidad siempre inacabada, por
ser indefinible la verdad con un cerebro finito y un universo de estructuras de
información infinitas. Traducir nuestra experiencia científica palabra-mundo, es la tarea de
composición escrita, misma que
caracteriza la actividad de investigación científica y las prácticas
disciplinares de corte científico. Pero nunca nuestra meta real es la verdad, sino
la realidad, producto de un consenso dinámico dentro de las diferentes escuelas
epistémicas de la ciencia. Puede ser algo extraño describir la actividad
científica de esta manera, pero es importante advertir al estudiante
universitario que la biología no es un asunto tecnológico y sin valores
morales, sino una actividad de composición escrita de correlación con la
realidad del hombre (objetividad). Sin embargo, el filósofo alemán Jürgen Habermas insiste en que no
hay nada en la noción de objetividad salvo la noción de acuerdo intersubjetivo: es el acuerdo alcanzado con interlocutores en la
literatura científica original, grupos de revisión o pares de trabajo, mediante
discusión libre y abierta de todas las hipótesis, alcanzando la hipótesis más
útil dentro de las políticas de verdad de su momento (criterios de verdad).
Esto último, debería ser un marco formativo del estudiante universitario de
ciencias, que sigue atrapado aún hoy en México en la idea de que instrumentos
de laboratorio pueden observar sin error y determinar la realidad, que en
muchos casos para empeorar las cosas ésta última se confunde con la verdad.
Campos
disciplinares de la nueva biología.
La nueva organización de la biología para su estudio, se da bajo el
paradigma del estudio de sistemas biológicos, a partir de la primera década del
siglo XXI se reconforman en varios contextos, entre los más relevantes
destacan:
Parafraseando al prominente biólogo Eileen E. M. Furlong
[71]
: La biología
de sistemas ha tenido un
crecimiento explosivo en el número de personas que participan en este ámbito de
la investigación y el número de publicaciones sobre el tema. Y, sin embargo,
los paradigmas que subyacen en el campo no han visto una expansividad similar a los que generaron los clásicos como Mendel o Darwin. En cambio, la
mayoría de los paradigmas que introducen, se han tomado de otros campos como la
ingeniería, la física y las matemáticas. Como resultado, un pequeño conjunto de
conceptos dominan el campo. El biólogo tradicional es visto por muchos como
anticuado y tolerado sólo como una fuente de datos. En este punto de vista, las ideas del biólogo
pueden incluso ser consideradas irrelevantes conceptual y teóricamente. En esta
perspectiva, uno mira críticamente a algunos de los paradigmas de la biología
de sistemas y pregunta si las ideas del biólogo, métodos y teorías ¿realmente
han quedado desfasadas? Vemos el futuro
de la biología de sistemas como una estrecha conexión entre los métodos in vivo e in vitro de la bioingeniería con modelado in silicón multicelulares y la biología sintética. Sin embargo, tome en cuenta que la
literatura muestra un Efecto Mateo provocado por los intereses de la industria
farmacéutica y biotecnológica que son la fuente de financiamiento principal de
la biología de sistemas, en general, parece más importarles las patentes y
otros registros industriales que la caza de la realidad científica.
Apoyándonos en el trabajo de Marc Vidal y Eileen E. M. Furlong, que en 2004 modelan la biología de
sistemas, intentaremos brevemente describir la nueva organización de la
biología moderna, ver figura 5.
Fig. 5. Biología de
sistemas
[72]
.
Para dar un ejemplo de los nuevos desafíos de la biología, en particular
la de sistemas, citamos un artículo de la prestigiosa revista británica de MacMillan Nature,
Molecular Systems Biology
[73]
:
El diseño y síntesis de circuitos básicos funcionales son las tareas
fundamentales de la biología sintética. Antes de que sea posible ingeniería de
orden superior, redes genéticas que pueden realizar funciones complejas, un
conjunto de herramientas de los dispositivos de base deben ser desarrolladas.
Entre esos dispositivos, circuitos lógicos secuenciales se espera que sean el
fundamento de los sistemas de procesamiento de la información genética. En este
estudio, se presenta el diseño y la construcción de un circuito lógico
secuencial genético en Escherichia coli. (Véase
fig. 6) Puede generar resultados
diferentes en respuesta a la misma señal de entrada sobre la base de su estado
interno, y "memorizar" las salidas. El circuito se compone de dos
partes: (1) un módulo de conmutación de memoria biestable y (2) un promotor de doble módulo NI, puerta reprimida. Los dos módulos fueron
diseñados de forma individual, y se acoplan entre sí por el ajuste de las
piezas de interconexión a través de la evolución dirigida. Después de puesta a
punto, el circuito podría ser varias veces, en su defecto provocado por la
misma señal de entrada, sino que funciona como un pulsador en el interruptor de
empuje.
Fig. 6. Circuito lógico de secuencia genética.
Fuente: Chunbo Lou, (2010)
Synthesizing a novel genetic sequential logic circuit: a push-on push-off
switch Molecular Systems Biology 6 (350) http://www.nature.com/msb/journal/v6/n1/full/msb20102.html
a)
Biología Cuántica.(Quantum Biology)
La física
cuántica parece la parte de la física más alejada de la biología, ya que la
coherencia cuántica parece poco importante en macromoléculas bioquímicas. Sin
embargo, el estudio de la fotosíntesis en algas indica que su alta eficiencia
es debida al uso de la coherencia cuántica. Por primera vez, dicho fenómeno ha
sido observado experimentalmente a temperatura ambiente (antes se había
observado por debajo de 77 K). Las proteínas fotosintéticas que absorben fotones
solares y excitan electrones en moléculas de clorofila actúan como un
computador cuántico (véase fig. 7).
Fig. 7. Fotosíntesis a temperatura ambiente.
Fuente: Elisabetta Collini, Cathy Y.
Wong, Krystyna E. Wilk,
Paul M. G. Curmi, Paul Brumer & Gregory D. Scholes (2009) Coherently wired light-harvesting in
photosynthetic marine algae at ambient temperature Nature 463, 644-647 http://www.nature.com/nature/journal/v463/n7281/full/nature08811.html
b)
Biología
Genómica/epigenómica (Genomics/epigenomicsBiology)
Lectura
de la información del genoma (bioinformática) depende de la interacción
compleja de cómo el ADN está empaquetado por proteínas. Una nueva era se abre
para los biólogos que participan en la comprensión de los sistemas celulares.
Describen el tipo de conocimientos sin precedentes que están surgiendo de las
investigaciones de cómo un único genoma de los mamíferos puede ser regulado
para producir diferentes tipos de células.
Fuente: Tarjei S. Mikkelsen, et al. (2007) Genome-wide maps of chromatin state in pluripotent and
lineage-committed cells. Nature 448, 553-560
http://www.nature.com/nature/journal/v448/n7153/full/nature06008.html
c)
Biología
Transcriptómica (Trancriptomics biology)
Advenimiento
de la genómica funcional ha permitido a las biociencias moleculares un largo
camino hacia la caracterización de los componentes moleculares de la vida. Sin
embargo, el reto para la biología en general es entender cómo funcionan los
organismos. Al descubrir cómo se plantea en función de las interacciones
dinámicas, biología de sistemas se refiere a la carencia de conocimiento entre
las moléculas y la fisiología. Los mejores sistemas establecen la biología
molecular que identifica las redes de interacción sobre la base del
comportamiento molecular, correlación observada en todos los estudios “ómicos". La biología de los sistemas de fondo examina
los mecanismos mediante los cuales las propiedades funcionales surgen en las
interacciones de los componentes conocidos de los principios que sustentan la
función celular. RNA-Seq es un enfoque desarrollado
recientemente para transcriptoma de perfiles que
utiliza tecnologías de secuenciación de profundidad. Los estudios que utilizan
este método ya han alterado nuestro punto de vista de la magnitud y la
complejidad de transcriptomas eucariotas. RNA-Seq también proporciona una medición mucho más precisa que
otros métodos de los niveles de transcripción y de sus isoformas.
Fuente: Zhong Wang, Mark
Gerstein & Michael Snyder (2009) RNA-Seq: a
revolutionary tool for transcriptomics. Nature
Reviews Genetics 10, 57-63 http://www.nature.com/nrg/journal/v10/n1/full/nrg2484.html
d)
Biología
Proteómica (Proteomic biology)
Los
éxitos recientes ilustran el papel de la masa proteómica, la espectrometría de
masas es una herramienta indispensable para la biología molecular, celular y
para el emergente campo de la biología de sistemas. Estos incluyen el estudio
de las interacciones proteína-proteína a través de aislamientos de afinidad
basada en una escala pequeña y grande de proteoma, la asignación de los
orgánulos numerosos, la descripción concurrentes del genoma y el proteoma
parásito de la malaria, y la generación de perfiles de proteínas cuantitativas
de diversas especies. La capacidad de la espectrometría de masas para
identificar y, cada vez más, para cuantificar con precisión miles de proteínas
de muestras complejas se puede esperar un impacto en términos generales en la
biología y la medicina.
Fuente: Benjamin F. Cravatt, Gabriel M. Simon & John R. Yates III (2007)
Review Article The biological impact of mass-spectrometry-based proteomics
Nature 450, 991-1000
http://www.nature.com/nature/journal/v450/n7172/full/nature06525.html
e)
Biología
de Redes de trabajo (Network biology)
Comprender
la organización funcional de la célula sistemáticamente, Catálogo de todas las
moléculas y sus interacciones dentro de una célula viva. Hay una clara
necesidad de entender cómo estas moléculas y las interacciones entre ellos
determinar la función de esta maquinaria de enorme complejidad, tanto de forma
aislada y cuando están rodeadas por otras células. Los rápidos avances en la
biología de la red indican que las redes celulares se rigen por leyes
universales y ofrecen un nuevo marco conceptual que podría revolucionar nuestra
visión de la biología y la patología de la enfermedad en el siglo XXI.
Fuente: Piano, F., et al. (2006) C. elegans network
biology: a beginning. WormBook, ed. The C. elegans Research. http://www.wormbook.org/chapters/www_networkbio/networkbio.html
f)
Biología Metabolómica (Metabolomics Biology)
Las nuevas técnicas para
la adquisición de datos metabolómicos siguen apareciendo. Esos datos requieren
condiciones adecuadas de almacenamiento en bases de datos correctamente
configuradas, el cual permite determinar el nivel de metabolomas microbianos (cientos de metabolitos principales)
y permitir que la naturaleza, organización y control de las redes metabólicas
se investigue. Una variedad de algoritmos para la reconstrucción de la red
metabólica acoplados a modelar algoritmos, son el sustantivo para el desarrollo
de la red metabólica y biología de sistemas. Incluso modelos cualitativos de
las redes metabólicas, al ser sometidos a las limitaciones estequiométricas,
puede ser muy informativo, y son el primer paso a los modelos cuantitativos, la
única que puede permitir la verdadera representación de los sistemas
bioquímicos complejos. A diferencia de las vías
de señalización, las redes metabólicas están sujetas a estrictas limitaciones estequiométricas. La metabolómica amplifica los cambios en el proteoma, y
representa más de cerca el fenotipo de un organismo.
Recientes avances permiten la producción (y codificación legible por ordenador
como SBML) de modelos de redes metabólicas reconstruyendo a partir de
secuencias del genoma, así como las medidas experimentales de gran parte del
metaboloma.
Fuente: Mo ML, Palsson (2009) Understanding human metabolic physiology: a
genome-to-systems approach. Trends Biotechnol. 27(1):37-44. http://www.ncbi.nlm.nih.gov/pubmed/19010556
g)
Biología Fenómica (Phenomics Biology)
Con
los avances en las tecnologías de genotipo de alto rendimiento, el paso
limitante de la velocidad de las investigaciones genéticas a gran escala se ha
convertido en la colección del fenotipo sensible y específico, la información
en una amplia muestra de participantes. Los médicos creen que desempeña un
papel fundamental para el éxito de los estudios de genética porque la clínica
puede aumentar sustancialmente el poder de estudio mediante la reducción de
errores de medición y mejorar la precisión diagnóstica para la investigación.
Fenómica es la medición y el análisis sistemático de los rasgos cualitativos y
cuantitativos, incluyendo la evolución clínica, y los métodos de imagen, para
el perfeccionamiento y la caracterización de un fenotipo. La fenómica requiere
fenotipificación de profundidad, la percepción de una amplia base de fenotipos
con la resolución y análisis de la Fenómica, integrado por la construcción de
mapas de calor, análisis de conglomerados, la minería de textos y análisis de rutas.
En el pasado, los biólogos han caracterizado a las respuestas de una amplia
gama de especies de plantas a su entorno. Como resultado, los datos fenotípicos
de cientos de experimentos están ahora a disposición del público.
Lamentablemente, esta información no está estructurada de una manera que
permita el análisis cuantitativo y comparativo. Nuestro objetivo es llenar este
vacío mediante la construcción de una gran base de datos que actualmente
contiene los datos de los experimentos en 1000 y 800 especies. El enfoque
actual, que nos referimos como "meta-phenomics”,
representa un valioso instrumento para comprender la respuesta integrada de las
plantas al medio ambiente y podrían servir de referencia para los futuros
esfuerzos de fenotipificación, así como para la modelización de los efectos del
cambio global en tanto especies silvestres y cultivos.
Fuente: Poorter H, Niinemets U,
Walter A, Fiorani F, Schurr U. (2010) Un método para construir curvas de dosis-respuesta para una amplia
gama de factores ambientales y características de las plantas por medio de un
meta-análisis de los datos fenotípicos. J Exp Bot. [Epub ahead of
print] http://www.ncbi.nlm.nih.gov/sites/entrez
Constrúyase
un documento de revisión corta, viviendo los rigores intelectuales de su
composición escrita, formato de comunicación bajo alguna norma de estilo
editorial y cuidando la calidad de las fuentes de información (vigencia,
relevancia, factor de impacto y pertinencia) y su terminología especializada. Y
prepare una exposición argumentativa para defender su trabajo, bajo el esquema
intelectual llamado discusión.
-

Fig. 8. !Los jóvenes requieren una oportunidad para
conocer la ciencia de la vida más allá de los límites de sus profesores y sus
notas!
Ácidos nucleicos, Acumulación de mutaciones, Alelos, Alométrica, Ambientes Cantgotu, Argumentos, Argumentos antrópicos, Autocatalítica, Bacterias,
Bioinformación, Biología
cuántica, Biología de Redes de trabajo, Biología de sistemas, Biología fenómica,
Biología genómica, Biología metabolómica, Biología proteómica, Biología
sintética, Biología transcriptómica, Biomasa, Biósfera, Células HeLa, Cell Cycle Models,
Coherencia matemática, Complejidad, Compuestos químicos, Concepto científico, Concepto
filosófico, Desafío cognitivo, Dicotomía, Distribución de densidad, Distribución estadística, Ecuación de réplica, Ecuaciones
de Poisson, Entropía, Epistemología, Epítopes
conformacionales, Equilibrio puntuado, Equilibrio químico, Estabilidad mecánica,
Estado físico, Estequiometria, Evolución, Factor de impacto, Fenómeno discreto, Fenomenología, Fenómica, Formas
caóticas, Genómica, Grados de libertad, Incertidumbre, Información, Interdisciplinariedad,
Investigación científica, Materia inorgánica, Mecánica cuántica, Membrana
celular, Momento físico, Mutaciones, Objetividad, Pleiotropía
antagónica, Polipéptido, Postura filosófica, Potenciales químicos, Práctica
científica, Prebiótico, Precámbrico tardío, Problema científico, Producto químico, Proteínas, Proteómica,
Protistas, Quirales, Racionalista, Revistas indexadas, Robustez mutacional, Selección
natural, Sistema de conocimiento, Talento, Tamaño de muestra unidad, Teoría de
campos, Teoría de juegos, Theory Cell, Theory Life, Termodinámica, Tiempo cinemicrográfico,
Transcriptómica, Universo subalterno, Variable aleatoria, Vida,
Ácidos nucleicos
|
Macromoléculas que constituyen el material genético, expresado mediante secuencia de monómeros llamados nucleótidos y unidos mediante
enlaces fosfodiéster |
Alelos
|
Una de las formas
variantes de un gen en un locus (posición) o de un marcador particular en un
cromosoma |
Alométrica
|
Cambios de dimensión
relativa de las partes corporales correlacionados con los cambios en el
tamaño total |
|
|
Argumentos
|
Razonamiento
empleado para demostrar, explicar y persuadir , formado por más de dos
proposiciones |
Argumentos antrópicos
|
Principios que
establecen que cualquier teoría válida sobre el universo tiene que ser
consistente con la existencia del ser humano. En otras palabras: "Si
en el Universo se deben verificar ciertas condiciones para nuestra
existencia, dichas condiciones se verifican ya que nosotros existimos". |
Autocatalítica
|
Colección de
entidades donde cada una de estas puede ser creada catalíticamente por otras
entidades dentro del conjunto, de manera que el conjunto es capaz de
catalizar su propia producción. |
Bacterias
|
Microorganismos
unicelulares, procariotas, tamaño 0,5 - 5 μm de diversas formas, incluyendo esferas, barras y hélices. Generalmente poseen
una pared celular de peptidoglicano. Muchas
disponen de flagelos o de otros sistemas de desplazamiento |
Bioinformación
|
Campo de informática
y la teoría de la información geométrica y simbólica de la vida |
Biología cuántica
|
Campos de
investigación... [donde]... se trata de problemas biológicos para cuyo
análisis es menester acudir a la física cuántica" |
Biología de sistemas
|
Campo de
investigación interdisciplinaria de los procesos biológicos en el que las
interacciones de los elementos, internos y externos, que influyen en el
desarrollo del proceso se representan con un sistema matemático |
Biología metabolómica
|
Conjunto de ciencias
y técnicas dedicadas al estudio completo del sistema biológico constituido por moléculas, intermediarios
metabólicos, metabolitos, hormonas y otras moléculas señal |
Biología sintética
|
Reúne la
ingeniería y las ciencias de la vida para diseñar y construir nuevos insumos
biológicos que no existen en el mundo natural (organismos y artefactos) o
para modificar los diseños existentes en los sistemas biológicos |
Biomasa
|
Materia total de los
seres que viven en un lugar determinado, expresada en peso por unidad de área
o de volumen |
Biósfera
|
Sistema material
formado por el conjunto de los seres vivos propios del planeta Tierra, junto
con el medio físico que les rodea y que ellos contribuyen a conformar |
Células HeLa
|
Línea de células epiteliares humanas procedentes de un carcinoma cervical,
primeras células humanas de las cuales se estableció una línea celular
permanente |
Coherencia matemática
|
Rigor de lógico
|
Complejidad
|
El grado mayor en el
que se puede mantener la coherencia de argumentos formado por términos no
lógicos y operadores lógicos.
|
Compuestos químicos
|
Sustancia formada
por la unión de dos o más elementos de la tabla periódica, y tiene fórmula
química
|
Concepto científico
|
Cualquier
conocimiento susceptible de una evolución intersubjetiva de su grado de
verdad sobre cualquier porción del
universo verificado completamente o parcialmente
|
Concepto filosófico
|
Instrumento de
exploración de la realidad, renovado sistemáticamente por la actividad del
pensar y que sirve para delinear los paradigmas de las escuelas del
pensamiento
|
Dicotomía
|
Fenómeno que implica
la separación en dos elementos o partes, especialmente cuando son opuestos:
bipartición |
Distribución estadística
|
Función que asigna a
cada suceso definido sobre la variable aleatoria la probabilidad de que dicho
suceso ocurra |
Ecuaciones de Poisson
|
Ecuación en
derivadas parciales con una amplia utilidad en electrostática, ingeniería
mecánica y física teórica. Aplicada en el estudio de las macromoléculas
describe el equilibrio
estático de una membrana ,esto es: esta ecuación gobierna la deflexión de la membrana sometida a tensiones
internas caracterizadas por el
parámetro bajo la acción de la carga |
Epistemología
|
Rama de la filosofía
cuyo objeto de estudio es el conocimiento: circunstancias históricas,
psicológicas y sociológicas que llevan a su obtención, y criterios por los
cuales se justifica o invalida |
Epítopes conformacionales
|
El epítope o determinante antigénico es la región de una
proteína o antígeno que es reconocida por un anticuerpo y que se une a él
para formar el complejo antígeno-anticuerpo. Están constituidos por
aminoácidos que, aunque están alejados
en la secuencia primaria de la proteína, se aproximan cuando esta se pliega
para lograr su estructura tridimensional. |
Equilibrio puntuado
|
Es
una teoría en biología evolutiva que propone que en la mayoría de la
reproducción sexual, las especies experimentaran pocos cambios evolutivos para la mayoría de
su historia geológica, permaneciendo en un estado extendido de inmovilización |
Equilibrio químico
|
El estado en el que
las actividades químicas o las concentraciones de los reactivos y los
productos no tienen ningún cambio neto en el tiempo. Este sería el estado que
se produce cuando el proceso químico evoluciona hacia adelante en la misma
proporción que su reacción inversa. |
Estequiometría
|
Rama
de la química que se ocupa de calcular la relación entre las cantidades de
sustancias que intervienen en un equilibrio de las reacciones químicas de
reacción o que combinan para formar un compuesto químico |
Fenomenología
|
Es
una meditación lógica que pretende superar las propias incertidumbres de la
lógica, orientándose hacia y con un lenguaje o logos que excluya la
incertidumbre entre la relación que hay entre los hechos (fenómenos) y el
ámbito en que se hace presente esta realidad |
Fenómica
|
Es el estudio de la
naturaleza de los fenotipos y la forma en que se determinan, en particular
cuando se estudia en relación con el conjunto de todos los genes |
Membrana celular
|
Estructura laminar
que engloba a las células, define sus límites y contribuye a mantener el
equilibrio entre el interior (medio intracelular) y el exterior (medio
extracelular) de éstas. Compuesta por una lámina que sirve de
"contenedor" para el citosol y los distintos compartimentos
internos de la célula, así como también otorga protección mecánica. Está
formada principalmente por fosfolípidos, colesterol, glúcidos y proteínas. |
Mutaciones
|
Es una alteración o
cambio en la información genética (genotipo) de un ser vivo que produce un cambio de características, que
se presenta súbita y espontáneamente, y que se puede transmitir o heredar a
la descendencia. |
Objetividad
|
Imparcialidad
con que se trata o se considera un asunto prescindiendo de las consideraciones
y los criterios personales o subjetivos. |
Pleiotropía antagónica
|
Teoría mantenida
durante largo tiempo sobre la causa de senectud considerada como el declive
de la fuerza de selección natural como una función de la edad de las células
somáticas adultas |
Polipéptido
|
Nombre utilizado
para designar un péptido de tamaño suficientemente grande; como orientación formado por una cadena de entre 10 y 50 aminoácidos |
Postura filosófica
|
Definición teórica y
práctica frente a una interrogación de la realidad |
Potenciales químicos
|
Es la variación en una función de estado
termodinámica característica por la variación en el número de moléculas.
Dependiendo de las condiciones experimentales, la función de estado
termodinámica característica es o bien la energía interna, la entalpía,
la energía libre de Gibbs, o la energía
libre de Helmholtz. |
Precámbrico tardío
|
Es la segunda
división geológica del Tiempo Precámbrico. Comienza hace 3.800 millones de
años (después del Eón Hadeico) y finaliza hace
2.500 millones de años (cuando comienza el Eón Proterozoico) durando 1300
millones de años. |
Producto químico
|
Es un conjunto de
compuestos químicos (aunque en ocasiones sea uno solo) destinado a cumplir
una función. Generalmente el que cumple la función principal es un solo
componente, llamado componente activo |
Proteínas
|
Son
macromoléculas formadas por la unión de varios aminoácidos, unidos mediante
enlaces peptídicos, están compuestas por carbono, hidrógeno, oxígeno
y nitrógeno. La mayoría también contienen azufre y fósforo. El orden y
disposición de los aminoácidos en una proteína depende del código genético.
|
Proteómica
|
Es
el estudio y caracterización de todo el conjunto de proteínas expresadas de
un genoma (proteoma), permite identificar, categorizar y clasificar las
proteínas con respecto a su función y a las interacciones que establecen
entre ellas. De este modo, se pueden caracterizar las redes funcionales que
establecen las proteínas y su dinámica durante procesos fisiológicos y
patológicos. |
Protistas
|
Son aquellos
organismos eucariontes que no pueden clasificarse dentro de alguno de los
otros tres reinos eucarióticos: Fungi,
Animalia o Plantae. Se desarrollan en ambientes
terrestres húmedos o en el medio interno de otros organismos. Pueden ser
unicelulares o pluricelulares, están dotadas de movilidad por reptación o por
cilios y flagelos, son autótrofos o heterótrofos. Algunos presentan
simultáneamente los dos modos de nutrición. Son de origen aerobios y algunos son
secundariamente anaerobios, tras haberse adaptado. |
Quirales
|
Isómeros que son
imágenes especulares (una es la imagen en el espejo de la otra) se llaman enantiómeros. Estas moléculas no son superponibles con
sus imágenes especulares, carecen de ejes de rotación |
Racionalista
|
Que coloca la razón
por encima de los sentimientos y las emociones, y asume que todo el universo
es posible de ser racionalizado |
Revistas indexadas
|
Son
aquellas publicaciones periódicas de carácter científico y tecnológico
contenidas en las bases de datos y que han pasado por un proceso de
selección y análisis por parte de las instituciones o empresas documentarias
que se realizan. |
Selección natural
|
Es un mecanismo
evolutivo que se define como la reproducción diferencial de los genotipos en
el seno de una población biológica. La formulación clásica de la selección
natural establece que las condiciones de un medio ambiente favorecen o
dificultan, es decir, seleccionan la reproducción de los organismos vivos
según sean sus peculiaridades |
Sistema de conocimiento
|
Organización
coherente de instrumentos epistemológicos que gestión la información y la
transforman en representaciones hipotéticas de la realidad |
Talento
|
Es una manifestación
de la inteligencia emocional y es una aptitud o conjunto de aptitudes o
destrezas para realizar una tarea determinada en forma exitosa |
Teoría de campos
|
Es el conjunto de
principios y técnicas matemáticas que estudia la dinámica y distribución
espacial de un campo físico |
Teoría de juegos
|
Es un área de la
matemática aplicada que utiliza modelos para estudiar interacciones en
estructuras formalizadas de incentivos (los llamados juegos) y llevar
a cabo procesos de decisión |
Termodinámica
|
Es una rama de la
física que estudia los efectos de los cambios de magnitudes de los sistemas a
un nivel macroscópico. Constituye una teoría fenomenológica, a partir de
razonamientos deductivos, que estudia sistemas reales, sin modelizar y sigue un método experimental[.] Los cambios estudiados son los de temperatura, presión y volumen, aunque
también estudia cambios en magnitudes como la imanación, el potencial
químico, la fuerza electromotriz, la gravedad y el estudio de los medios
continuos en general |
Transcriptómica
|
La transcriptómica
estudia y compara transcriptomas, es decir, los conjuntos de ARN mensajeros o
transcriptos presentes en una célula, tejido u organismo. La transcriptómica
se caracteriza por el monitoreo y análisis de expresión simultáneo de muchos
genes utilizando macro- o micro-matrices de ADN (microarrays o chips de ADN). |
Variable aleatoria
|
Es una función real
definida en el espacio muestral asociado a un
experimento aleatorio, Ω. También
se le llama variable de azar o variable estocástica, y significa cantidad que
puede tomar varios valores imprevistos. |
Ésta fue publicada originalmente en Internet bajo la categoría de contenido abierto sobre la URL: http://dieumsnh.qfb.umich.mx/ mismo título y versión de contenido digital. Este es un trabajo de autoría publicado sobre Internet Copyright © 2003-2010 por la Coordinación de Innovación Educativa Q.F.B./UMSNH, protegido por las leyes de derechos de propiedad de los Estados Unidos Mexicanos. No puede ser reproducido, copiado, publicado, prestado a otras personas o entidades sin el permiso explícito por escrito de la ANUIES/SINED/UMSNH o por los Autores.
Ochoa, H. E., et al (2010) Biología de sistemas. Una mirada a lo más profundo de nuestro origen, evolución y funcionamiento. México, ANUIES/SINED/UMSNH. vii, 302 p.; carta




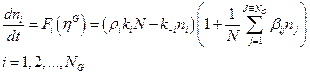
{kind=link}